Sulforaphane is a phytochemical, a substance within the isothiocyanate group of organosulfur compounds, found in cruciferous vegetables, such as broccoli, cabbage, cauliflower, and Brussels sprouts. It can also be found in bok choy, kale, collards, mustard greens and watercress. Research studies have shown that sulforaphane can help prevent various types of cancer by activating the production of Nrf2, or nuclear factor erythroid 2-related factor, a transcription factor which regulates�protective antioxidant mechanisms that control the cell’s response to oxidants. The purpose of the following article is to describe the function of sulforaphane.
Abstract
The KEAP1-Nrf2-ARE antioxidant system is a principal means by which cells respond to oxidative and xenobiotic stresses. Sulforaphane (SFN), an electrophilic isothiocyanate derived from cruciferous vegetables, activates the KEAP1-Nrf2-ARE pathway and has become a molecule-of-interest in the treatment of diseases in which chronic oxidative stress plays a major etiological role. We demonstrate here that the mitochondria of cultured, human retinal pigment epithelial (RPE-1) cells treated with SFN undergo hyperfusion that is independent of both Nrf2 and its cytoplasmic inhibitor KEAP1. Mitochondrial fusion has been reported to be cytoprotective by inhibiting pore formation in mitochondria during apoptosis, and consistent with this, we show Nrf2-independent, cytoprotection of SFN-treated cells exposed to the apoptosis-inducer, staurosporine. Mechanistically, SFN mitigates the recruitment and/or retention of the soluble fission factor Drp1 to mitochondria and to peroxisomes but does not affect overall Drp1 abundance. These data demonstrate that the beneficial properties of SFN extend beyond the activation of the KEAP1-Nrf2-ARE system and warrant further interrogation given the current use of this agent in multiple clinical trials.
Sulforaphane is an Nrf2-Independent Inhibitor of Mitochondrial Fission
Sulforaphane (SFN) is an isothiocyanate compound derived in the diet most commonly from cruciferous vegetables [56]. It is generated in plants as a xenobiotic response to predation via vesicular release of the hydrolytic enzyme myrosinase from damaged cells; this enzyme converts glucosinolates to isothiocyantes [42]. Over the last two decades, SFN has been extensively characterized for its reported anticancer, antioxidant, and antimicrobial properties [57]. Much of this efficacy has been attributed to the capacity of SFN to modulate the KEAP1-Nrf2-antioxidant response element (ARE) signaling pathway, although additional activities of the compound have been identified, including the inhibition of histone deacetylase activity and cell cycle progression [29]. Nrf2 is the master antioxidant transcription factor and under conditions of homeostasis, its stability is suppressed through the action of the cytoplasmic Cullin3KEAP1 ubiquitin ligase complex [20]. Specifically, Nrf2 is recruited to the Cullin3KEAP1 ligase by binding to the dimeric substrate adaptor KEAP1 and is subsequently modified with polyUb chains that target the transcription factor for proteasome-mediated degradation. This constitutive turnover limits the half-life of Nrf2 in unstressed cells to ~15 min [30], [33], [46], [55]. In response to numerous types of stress, most notably oxidative stress, KEAP1, a cysteine-rich protein, acts as a redox sensor, and oxidative modification of critical cysteines, particularly C151, of KEAP1 dissociates Nrf2-KEAP1 from CUL3 thereby preventing Nrf2 degradation [8], [20], [55]. Notably, SFN, and possibly other Nrf2 activators, mimic oxidative stress by modifying C151 of KEAP1 e.g. [21]. Stabilization of Nrf2 allows for its translocation to the nucleus where it induces the expression of a battery of Phase II antioxidant and detoxification genes. Nrf2 binds to the antioxidant response promoter elements (ARE) of its cognate target genes through heterodimerization with small Maf proteins [19]. This system presents a dynamic and sensitive response to indirect antioxidants like SFN, free radicals generated by the mitochondria [16], or other physiologic sources of oxidative stress [41].
Mitochondria are dynamic, subcellular organelles that regulate a host of cellular functions ranging from ATP production and intracellular calcium buffering to redox regulation and apoptosis [13], [49]. Mitochondria also represent the principal source of reactive oxygen species (ROS) within the cell. Proper regulation of mitochondrial function is therefore necessary for optimizing ATP production to meet cellular needs while simultaneously minimizing the potentially harmful effects of excessive free radical production. A critical requirement for fine modulation of mitochondrial function is the capacity for mitochondria to function both independently as biochemical machines and as part of a vast, responsive network.
Mitochondrial network morphology and function are determined by a regulated balance between fission and fusion. Mitochondrial fission is required for daughter cell inheritance of mitochondria during cell division [28] as well as for the selective, autophagic degradation of depolarized or damaged mitochondria, termed mitophagy [1]. Conversely, fusion is required for complementation of mitochondrial genomes and sharing of electron transport chain components between neighboring mitochondria [54]. At the molecular level, mitochondrial fission and fusion are regulated by large, dynamin-like GTPases. Three enzymes primarily regulate fusion: Mitofusins 1 and 2 (Mfn1/2) are two-pass outer membrane proteins that mediate outer membrane fusion via heterotypic interactions between adjacent mitochondria [15], [25], [37], while OPA1 is an inner membrane protein that simultaneously ensures matrix connectivity by regulating the melding of inner membranes [5]. The GTPase activity of all three proteins is required for robust fusion [5], [18], and OPA1 is further regulated by complex proteolysis within the mitochondrial inner membrane by the proteases OMA1 [14], PARL [6], and YME1L [45]. Importantly, intact mitochondrial membrane potential is required for efficient fusion in order to suppress integration of damaged and healthy mitochondria [26].
Mitochondrial fission is primarily catalyzed by a cytosolic protein called Dynamin-related protein 1 (Drp1/DNM1L). Drp1 is recruited from the cytosol to prospective sites of fission on the mitochondrial outer membrane [43]. The major receptors for Drp1 on the outer membrane are mitochondrial fission factor (Mff) [32] and, to a lesser extent, Fission 1 (Fis1) [51]. Additionally, a decoy receptor, MIEF1/MiD51, was discovered that acts to further limit the activity of Drp1 protein at potential fission sites [58]. Once docked at the mitochondrial outer membrane, Drp1 oligomerizes into spiral-like structures around the body of the mitochondrion and then utilizes the energy derived from GTP hydrolysis to mediate the physical scission of the mitochondrial outer and inner membranes [17]. Endoplasmic reticulum-derived tubules act as an initial constrictor of mitochondria prior to Drp1 oligomerization, underscoring the revelation that non-constricted mitochondria are wider than the permissive circumference of a completed Drp1 spiral [12]. Actin dynamics are also important for the ER-mitochondria interactions that precede mitochondrial fission [24]. In addition to its role in mitochondrial fission, Drp1 catalyzes the fission of peroxisomes [40].
Drp1 is very similar to the well-characterized dynamin protein in that both proteins contain an N-terminal GTPase domain, a Middle domain that is critical for self-oligomerization, and a C-terminal GTPase effector domain [31]. Drp1 achieves selectivity for mitochondrial membranes through a combination of interactions with its receptor proteins Mff and Fis1 and also through its affinity for the mitochondria-specific phospholipid cardiolipin via the unique B-insert domain of Drp1 [2]. Drp1 typically exists as a homotetramer in the cytoplasm, and higher order assembly at mitochondrial fission sites is mediated by the Middle domain of Drp1 [3].
Given the implicit link between mitochondrial function and the KEAP1-Nrf2-ARE pathway, we investigated the effects of Nrf2 activation on mitochondrial structure and function. We demonstrate here that SFN induces mitochondrial hyperfusion that, unexpectedly, is independent of both Nrf2 and KEAP1. This effect of SFN is through an inhibition of Drp1 function. We further demonstrate that SFN confers resistance to apoptosis that is Nrf2-independent and mimics that observed in cells depleted of Drp1. These data collectively indicate that in addition to stabilizing and activating Nrf2, SFN modulates mitochondrial dynamics and preserves cellular fitness and survival.
Results
Sulforaphane Induces Nrf2/KEAP1-Independent Hyperfusion of Mitochondria
In the course of studying the effects of Nrf2 activation on mitochondrial network dynamics, we discovered that treatment of immortalized, human retinal pigment epithelial (RPE-1) cells with sulforaphane (SFN), a potent activator of Nrf2 signaling, induced a robust fusion of the mitochondrial network when compared with vehicle-treated control cells (Fig. 1A and B). The morphology of the mitochondria in these cells greatly resembled that of the mitochondria in cells depleted by siRNA of endogenous Drp1, the principal mitochondrial fission factor (Fig. 1A). This result raised the intriguing idea that mitochondrial fission and fusion status responds directly to Nrf2 levels in the cell. However, stimulation of cells with other Nrf2 stabilizers and activators such as the proteasome inhibitor MG132, the pro-oxidant tBHQ, or knockdown of the Nrf2 inhibitor KEAP1 did not induce mitochondrial fusion (Fig. 1A and B). Stabilization of Nrf2 by these manipulations was confirmed by western blotting for endogenous Nrf2 (Fig. 1C). Furthermore, expression of Nrf2 was dispensable for SFN-induced mitochondrial fusion, as knockdown of endogenous Nrf2 with siRNA failed to counter this phenotype (Fig. 1D�F). Because SFN stimulates the KEAP1-Nrf2-ARE pathway by covalently modifying cysteine residues of KEAP1 [21], we knocked down KEAP1 to address whether SFN-induced mitochondrial hyperfusion is stimulated through a KEAP1-dependent, but Nrf2 independent pathway. However, depletion of KEAP1 also failed to abrogate SFN-induced mitochondrial fusion (Fig. 1G�I). In fact, SFN reversed the pro-fission morphology induced by depletion of KEAP1 (Fig. 1G, panel b versus panel d). These results indicate that SFN treatment causes mitochondrial fusion independent of the canonical KEAP1-Nrf2-ARE pathway and led us to interrogate whether SFN directly affects components of the mitochondrial fission or fusion machinery.
Figure 1 SFN induces Nrf2/KEAP1-independent mitochondrial fusion. (A) RPE-1 cells were transfected with the indicated siRNAs and 3 days later treated with DMSO or the Nrf2 activators SFN (50 ?M), MG132 (10 ?M), or tBHQ (100 ?M) for 4 h. Mitochondria (red) are labeled with an anti-Tom20 antibody, and nuclei (blue) are counterstained with DAPI. (B) Graph showing quantification of mitochondrial morphology scoring from (A). >50 cells per condition were evaluated in a blinded fashion. (C) Representative western blots from (A). (D) RPE-1 cells were transfected with 10 nM siRNA and 3 days later treated with SFN for 4 h prior to being fixed and stained as in (A). (E) Graph showing quantification of mitochondrial phenotype scoring from (D). >100 cells per condition were evaluated in a blinded fashion. (F) Representative western blots from (D). (G) Cells were transfected and treated as in (D) with siCON or siKEAP1. (H) Cells from (G) were scored as in (B) and (E) on the basis of mitochondrial morphology. (I) Representative western blots from (G). Data in (B), (E), and (H) were compiled from 3 independent experiments each and statistical significance was determined by two-tailed Student’s t-test. Error bars reflect +/- S.D. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article).
Sulforaphane Impairs the Mitochondrial Association of Drp1
Based on the finding that SFN-treatment induces mitochondrial hyperfusion, we reasoned that this phenotype was either a consequence of excessive fusion activity or an inhibition of fission activity. To discriminate between these two possibilities, we compared the morphology of peroxisomes in the presence and absence of SFN. Peroxisomes are similar to mitochondria in that they are dynamic organelles the shape and length of which are constantly in flux [44]. Peroxisomes contain both Fis1 and Mff in their outer membrane and, as a consequence, are targets for Drp1-mediated fission [22], [23]. However, peroxisomes do not utilize the fusion machinery of the mitochondrial network and consequently, do not undergo fusion [39]. Rather, peroxisomal fission is opposed by the lengthening of existing peroxisomes via de novo addition of membranes and proteins [44]. Because peroxisomes lack Mfn1/2 and OPA1, we reasoned that if SFN activates the fusion machinery rather than inhibiting the fission machinery, peroxisome length would not be affected. In vehicle-treated cells, peroxisomes are maintained as short, round, punctiform organelles (Fig. 2, panels b and d). However, SFN treatment increased peroxisome length by ~2-fold as compared to control cells (Fig. 2, panels f and h). Furthermore, many of the peroxisomes were pinched near the center, indicating a potential scission defect (Fig. 2, panel h, arrowheads). Likewise, peroxisomes in cells transfected with Drp1 siRNA were abnormally long (Fig. 2, panels j and l), confirming that Drp1 is required for peroxisomal fission and suggesting that SFN-treatment causes mitochondrial and peroxisomal phenotypes by disrupting the fission machinery.
Figure 2 SFN induces peroxisomal lengthening. (A) RPE-1 cells were transfected with 10 nM of the indicated siRNA and 3 days later treated with DMSO or 50 ?M SFN for 4 h. Peroxisomes (green) were labeled with an anti-PMP70 antibody, mitochondria with MitoTracker (red), and DNA counterstained with DAPI. Enlarged insets of peroxisomes are shown on the right (panels d, h, and l) to facilitate visualization of the changes in morphology induced by SFN and Drp1 depletion. Arrowheads highlight constriction points. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article).
We next determined how SFN restricts Drp1 function. Possibilities included reductions in expression levels, recruitment/retention at mitochondria, oligomerization, or enzymatic activity of the GTPase. A deficit in any one of these would result in reduced mitochondrial fission and hyperfusion. We did not detect reproducible changes in Drp1 protein levels after SFN-treatment (Figs. 1C and 3A), and therefore concluded that SFN does not alter Drp1 stability or expression, consistent with Drp1 having a half-life of >10 h [50] and our SFN treatments being of shorter duration. Next, we investigated whether SFN affected the recruitment or retention of Drp1 to mitochondria. Fractionation studies showed that SFN induced a loss of Drp1 from the mitochondrial fraction (Fig. 3A, lanes 7�8 and Fig. 3B). As reported previously [43], only a minor fraction of Drp1 (~3%) is associated with the mitochondrial network at any given time during steady state conditions with most of the enzyme residing in the cytoplasm (Fig. 3A, lanes 5�8). These fractionation data were confirmed using co-localization analysis which showed a ~40% reduction in mitochondria-localized, punctate Drp1 foci after SFN-treatment (Fig. 3C and D). Together, these data indicate that the mitochondrial fusion induced by SFN is, at least partially, due to the attenuated association of Drp1 with the mitochondria. Our data do not distinguish between whether SFN interferes with the mitochondrial recruitment versus the mitochondrial retention of Drp1, or both, as the analysis of endogenous Drp1 was not amenable to visualizing the GTPase by live-cell microscopy.
Figure 3 SFN causes a loss of Drp1 from the mitochondria. (A) Subcellular fractionation of RPE-1 cells following 4 h of DMSO or SFN. Whole-cell lysates (WCL), nuclear (Nuc), cytosolic (Cyto), and crude mitochondrial (Mito) fractions were resolved by SDS-PAGE and processed for western blotting with the indicated antibodies. The migration of molecular weight markers is indicated on the left. (B) Graphs showing densitometric quantification of Drp1 in the indicated fractions from (A). (C) RPE-1 cells were transfected with 10 nM siCON or siDrp1 and 3 days later treated with DMSO or SFN for 4 h. Drp1 (green) was visualized with an anti-Drp1 antibody, mitochondria with MitoTracker (red), and nuclei with DAPI (blue). (D) Automated co-localization analysis of Drp1 and MitoTracker signal from (C). Data in (B) and (D) were compiled from 3 and 5 independent experiments, respectively, and statistical significance was determined by two-tailed Student’s t-test. Error bars reflect +/- S.D and asterisks denote statistical significance. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article).
Sulforaphane Confers Protection Against Staurosportine-Induced Apoptosis Independent of Nrf2
Previous work has shown that mitochondrial fission is permissive in the formation of pores in the outer mitochondrial membrane generated by Bax/Bak during apoptosis [11]. Drp1 has been shown to be selectively recruited to mitochondria during apoptosis [11] and, consistent with this, fragmented mitochondria have been observed early in the process [27]. Conversely, inhibiting mitochondrial fission is thought to inhibit apoptosis by blocking the formation of the outer membrane pores that allow for cytochrome c release [53]. Accordingly, stimulating mitochondrial fusion delays the progression of apoptosis induced by compounds including staurosporine (STS) [14]. To determine whether SFN protects RPE-1 cells from STS-mediated apoptosis and if so, whether this requires Nrf2, we established an assay to readily induce poly ADP ribose polymerase (PARP) cleavage, a substrate of activated caspase-3 and definitive marker of apoptosis. Treatment of RPE-1 cells with 1 �M STS for 6 h only caused a very modest cleavage of PARP yet this was prevented by SFN co-treatment (e.g., Fig. 4A, lane 3 versus 4). To increase the robustness of this assay, we further sensitized cells to STS-induced apoptosis by pre-treating them with siRNA targeting the anti-apoptotic factor, Bcl-XL. This pretreatment reduced the expression of Bcl-XL and markedly promoted PARP cleavage as a function of time exposed to STS (Fig. 4B, compare lane 2 to lanes 4�10). Importantly, 2 h of pre-treatment with SFN mitigated PARP cleavage in cells exposed to STS (Fig. 4C, lane 3 versus 4 and lane 5 versus 6). Likewise, cells stably depleted of Nrf2 by CRISPR/Cas9 were comparably protected from STS toxicity by SFN pre-treatment (Fig. 4C, lane 11 versus 12 and lane 13 versus 14 and Fig. 4D). This protection was observed using both PARP cleavage (Fig. 4C and D) and cellular morphology (Fig. 4E) as readouts. The efficacy of Nrf2 depletion by CRISPR/Cas9 was confirmed by western blotting (Fig. 4C, Nrf2 blot). As predicted, depleting cells of Drp1, which also yields a hyperfusion phenotype (Fig. 1A), also blocked PARP cleavage in response to STS as compared to control cells incubated with SFN (Fig. 4F and G). Together, these findings are consistent with SFN conferring protection against apoptosis through its capacity to restrict Drp1 function, independent of the stabilization and activation of Nrf2.
Figure 4 The cytoprotective effects of SFN are independent of Nrf2 expression (A) RPE-1 cells were pre-treated with DMSO or 50 ?M SFN for 2 h prior to treatment with DMSO, 1 ?M staurosporine (STS), or 50 ?M etoposide for 6 h and were processed for anti-PARP western blotting. (B) RPE-1 cells were transfected with 2.5 nM siCON, 1 nM siBcl-XL, or 2.5 nM siBcl-XL and 3 days later were treated with DMSO or 1 ?M STS for 2, 4, or 6 h. Representative western blots are shown and the migration of molecular weight markers is indicated on the left. (C) CRISPR/Cas9-generated wild-type (Nrf2WT) and Nrf2 knockout (Nrf2KO) RPE-1 cells were transfected with 1 nM siBcl-XL and 3 days later were pre-treated with DMSO or 50 ?M SFN for 2 h. Subsequently, the cells were treated with 1 ?M STS for 2, 4, or 6 h. Representative western blots with the indicated antibodies are shown. (D) Quantification of cleaved PARP as a percentage of total PARP (cleaved+uncleaved) from 3 independent experiments. Importantly, the levels of cleaved PARP were comparable whether cells expressed Nrf2 or not, indicating that SFN protection from STS is independent of the transcription factor. (E) 20X phase-contrast images taken immediately prior to harvest of lysates from (C). Scale bar=65 �m. (F) Representative western blots demonstrating that depletion of Drp1 confers near-comparable protection from STS as SFN treatment. RPE-1 cells were transfected with 1 nM siBcl-XL and additionally transfected with either 10 nM siCON or 10 nM siDrp1. 3 days later, siCON cells were pre-treated with SFN as in (A) and (C) and then exposed to STS for 4 h prior to being harvested and processed for western blotting with the indicated antibodies. (G) Same as (D) for the data presented in (F) compiled from 3 independent experiments. Error bars reflect +/- S.E.M.
Discussion
We have discovered that SFN modulates mitochondrial fission/fusion dynamics independent of its effects on the KEAP1-Nrf2-ARE pathway. This is intriguing because of an assumed link between mitochondrial dysfunction and ROS production and the necessity of squelching mitochondria-derived free radicals through the activation of Nrf2. This additional functional impact of SFN is of potential importance given the more than 30 clinical trials currently underway testing SFN for the treatment of a variety of diseases including prostate cancer, obstructive pulmonary disease, and sickle cell disease [7], [10], [47].
Because SFN is an isothiocyanate [56] and it activates Nrf2 signaling by directly acylating critical KEAP1 cysteines to suppress Nrf2 degradation [21], it follows that SFN exerts its pro-fusion effects by modulating the activity of a fission or fusion factor via cysteine modification. Our data strongly support Drp1 being negatively regulated by SFN although whether the GTPase is a direct target of acylation remains to be elucidated. Despite this knowledge gap, the function of Drp1 is clearly being compromised by SFN as both mitochondria and peroxisomes become hyperfused in response to SFN treatment and these organelles share Drp1 for their respective scission events [38]. In addition, SFN decreases the amount of Drp1 that localizes and accumulates at mitochondria (Fig. 3). Because our experiments were done with all endogenous proteins, our detection of Drp1 at mitochondrial fission sites is under steady-state conditions, and consequently, we cannot distinguish between a recruitment versus a retention defect of the enzyme caused by SFN. Further, we cannot eliminate the possibility that SFN acylates a receptor at the mitochondria (Fis1 or Mff) to block Drp1 recruitment yet, we suspect that Drp1 is directly modified. Drp1 has nine cysteines, eight of which reside within the Middle Domain that is required for oligomerization [3], and one of which resides in the GTPase Effector Domain (GED) at the C-terminus of Drp1. Direct acylation of any of these cysteines could cause an activity defect in Drp1 and therefore underlie the effect of SFN on mitochondrial dynamics. Notably, prior work suggests that defects in oligomerization and catalytic activity can abrogate the retention of Drp1 at the mitochondria [52]. Cys644 in the GED domain is a particularly attractive target based on previous work showing that mutation of this cysteine phenocopies mutations that impair Drp1 GTPase activity [4] and that this particular cysteine is modified by thiol-reactive electrophiles [9]. Resolution of this outstanding question will require mass spectrometric validation.In summary, we have identified a novel, cytoprotective function for the clinically-relevant compound SFN. In addition to activating the master anti-oxidant transcription factor Nrf2, SFN promotes mitochondrial and peroxisomal fusion, and this effect is independent of Nrf2. The mechanism underlying this phenomenon involves a reduction in the function of the GTPase Drp1, the primary mediator of mitochondrial and peroxisomal fission. A major consequence of SFN-mediated mitochondrial fusion is that cells become resistant to the toxic effects of the apoptosis inducer staurosporine. This additional cytoprotective action of SFN could be of particular clinical utility in the numerous neurodegenerative diseases for which age is the leading risk factor (e.g., Parkinson’s Disease, Alzheimer’s Disease, Age-related Macular Degeneration) as these maladies have been associated with apoptosis and reduced levels and/or dysregulation of Nrf2 [35], [36], [48]. Together, these data demonstrate that the cytoprotective properties of SFN extend beyond activation of the KEAP1-Nrf2-ARE system and warrant further studies given the current use of this agent in multiple clinical trials.
Materials and Methods
Apoptosis Assays
Cells were seeded and transfected with siRNA as indicated below. The cells were pre-treated with 50 ?M sulforaphane for 2 h to induce mitochondrial fusion and were then treated with 1 ?M staurosporine to induce apoptosis. At the time of harvest, media was collected in individual tubes and subjected to high speed centrifugation to pellet apoptotic cells. This cell pellet was combined with adherent cells and solubilized in 2 times-concentrated Laemmli buffer. Samples were subjected to anti-PARP western blotting.
CRISPR/Cas9 Construct Generation
To create LentiCRISPR/eCas9 1.1, LentiCRISPR v2 (addgene #52961) was first cut with Age1 and BamH1. Next, SpCas9 from eSpCas9 1.1 (addgene #71814) was PCR amplified with Age1 and BamH1 overhangs using the following primers (Forward AGCGCACCGGTTCTAGAGCGCTGCCACCATGGACTATAAGGACCACGAC, Reverse AAGCGCGGATCCCTTTTTCTTTTTTGCCTGGCCGG) and ligated into the cut vector above. sgRNA sequences were determined by using Benchling.com. Parameters were set to target the coding sequence with the highest on-target and lowest off-target scores. The following sequences (targeting sequence underlined, hs sgNFE2L2#1 sense CACCGCGACGGAAAGAGTATGAGC, antisense AAACGCTCATACTCTTTCCGTCGC; hs sgNFE2L2#2 sense CACCGGTTTCTGACTGGATGTGCT, antisense AAACAGCACATCCAGTCAGAAACC; hs sgNFE2L2#3 sense CACCGGAGTAGTTGGCAGATCCAC, antisense AAACGTGGATCTGCCAACTACTCC) were annealed and ligated into BsmB1 cut LentiCRISPR/eCas9 1.1. Lentivirally infected RPE-1 cells were selected with puromycin and maintained as a pooled population. Knockout was confirmed by immunofluorescence and western blotting.
Cell Culture and Transfections
Human retinal pigment epithelial cells transformed with telomerase (RPE-1) (ATCC) were cultured in Dulbecco’s Modified Eagle Medium (DMEM) containing 1 g/L glucose supplemented with penicillin, streptomycin, 1X non-essential amino acid cocktail (Life Technologies), and 10% Fetal Bovine Serum (Life Technologies). For siRNA-transfections, 30,000�35,000 cells/mL were seeded overnight. Cells received 10 nM siRNA diluted in serum-free DMEM and combined with 0.3% Interferin transfection reagent (PolyPlus). For apoptosis sensitization, cells received 1 nM Bcl-XL siRNA. Cells were harvested 2�3 days post-transfection.
Chemicals, Antibodies, and siRNA Oligos
Antibodies against ?-tubulin (Cell Signaling), ?-tubulin (Sigma), Drp1 (BD Biosciences), KEAP1 (Proteintech), Lamin B1 (Abcam), PARP (Cell Signaling), PMP70 (Abcam), and Tom20 (BD Biosciences) were used at 1:1000 dilutions for western blotting and for immunofluorescence. In-house, anti-Nrf2 rabbit antibody was used at 1:2000 for western blotting [34], [59]. Sulforaphane (Sigma) and staurosporine (Tocris) were used at 50 ?M and 1 ?M respectively. siRNAs against Drp1 (Dharmacon), Nrf2 (Dharmacon), KEAP1 (Cell Signaling), and Bcl-XL (Cell Signaling) were used at 10 nM unless otherwise noted.
Immunofluorescence and in Vivo Labeling
Cells seeded on 18 mm glass coverslips were treated with vehicle or drug, fixed in 3.7% formaldehyde and then permeabilized in 0.2% Triton X-100/PBS on ice for 10 min. Primary antibodies were incubated in 3% bovine serum albumin (BSA) in PBS overnight at 4 �C. Following PBS washes, cells were incubated for 1 h in species-appropriate, Alexa488- or Alexa546-, conjugated secondary antibodies (diluted 1:1000) and 0.1 ?g/mL DAPI (Sigma) in 3% BSA/PBS. Mitochondria were visualized either by anti-Tom20 immunofluorescence or by incubating cells in 200 nM MitoTracker Red CMXRos (Molecular Probes, Inc.) in serum-free DMEM for 30 min at 37 �C prior to fixation.
Microscopy and Image Analysis
Immunofluorescence samples were viewed on an LSM710 Confocal microscope (Carl Zeiss). Micrographs were captured using 63X or 100X oil immersion objectives and images adjusted and enhanced using Adobe Photoshop CS6. Co-localization analysis was performed using Carl Zeiss LSM710 co-localization feature with thresholds manually set while blinded to the identity of the samples. Scale bars throughout, unless otherwise indicated, are 10 �m. Mitochondrial morphology was assessed by blinded scoring. If the mitochondria of a cell were maintained as multiple, round, discriminate puncta, the cell was scored as �fission�. If individual mitochondria were indistinguishable and the whole mitochondrial network appeared continuous, the cell was scored as �fusion�. All other cells, including those with clustering mitochondria, were scored as �intermediate�.
Subcellular Fractionations
RPE-1 cells were grown to confluence. Following a PBS wash, cells were subjected to centrifugation at 600�g for 10 min and resuspended in 600 ?L isolation buffer (210 mM Mannitol, 70 mM Sucrose, 5 mM MOPS, 1 mM EDTA pH 7.4+1 mM PMSF). The suspension was lysed 30 times in a Dounce homogenizer. A fraction of the homogenate was preserved as a �whole cell lysate.� The remainder was subjected to centrifugation at 800�g for 10 min to pellet nuclei. Supernatants were subjected to centrifugation at 1500�g for 10 min to clear remaining nuclei and unlysed cells. This supernatant was subjected to centrifugation at 15,000�g for 15 min to pellet mitochondria. The supernatant was preserved as the �cytosolic fraction�. The pellet was washed gently with PBS and resuspended in isolation buffer. The protein concentration of each fraction was measured by bicinchoninic acid (BCA) assay and equivalent amounts of protein were resolved by SDS-PAGE.
Western Blotting
Cells were washed in PBS and solubilized in 2 times concentrated Laemmli solubilizing buffer (100 mM Tris [pH 6.8], 2% SDS, 0.008% bromophenol blue, 2% 2-mercaptoethanol, 26.3% glycerol, and 0.001% Pyrinin Y). Lysates were boiled for 5 min prior to loading on sodium dodecyl sulfate (SDS) polyacrylamide gels. Proteins were transferred to nitrocellulose membranes and the membranes were blocked for 1 h in 5% Milk/TBST. Primary antibodies were diluted in 5% Milk/TBST and incubated with the blot overnight at 4 �C. Horseradish peroxidase (HRP)-conjugated secondary antibodies were diluted in 5% Milk/TBST. Blots were processed with enhanced chemiluminescence and densitometric quantifications were performed using ImageJ software.
Sulforaphane is a chemical from the isothiocyanate collection of organosulfur substances obtained from cruciferous vegetables, including broccoli, cabbage, cauliflower, kale, and collards, among others. Sulforaphane is produced when the enzyme myrosinase transforms glucoraphanin, a glucosinolate, into sulforaphane, also known as sulforaphane-glucosinolate. Broccoli sprouts and cauliflower have the highest concentration of glucoraphanin or the precursor to sulforaphane. Research studies have demonstrated that sulforaphane enhances the human body’s antioxidant capabilities to prevent various health issues. Dr. Alex Jimenez D.C., C.C.S.T. Insight
Sulforaphane and Its Effects on Cancer, Mortality, Aging, Brain and Behavior, Heart Disease & More
Isothiocyanates are some of the most important plant compounds you can get in your diet. In this video I make the most comprehensive case for them that has ever been made. Short attention span? Skip to your favorite topic by clicking one of the time points below. Full timeline below.
Key sections:
00:01:14 – Cancer and mortality
00:19:04 – Aging
00:26:30 – Brain and behavior
00:38:06 – Final recap
00:40:27 – Dose
Full timeline:
00:00:34 – Introduction of sulforaphane, a major focus of the video.
00:01:14 – Cruciferous vegetable consumption and reductions in all-cause mortality.
00:02:12 – Prostate cancer risk.
00:02:23 – Bladder cancer risk.
00:02:34 – Lung cancer in smokers risk.
00:02:48 – Breast cancer risk.
00:03:13 – Hypothetical: what if you already have cancer? (interventional)
00:03:35 – Plausible mechanism driving the cancer and mortality associative data.
00:04:38 – Sulforaphane and cancer.
00:05:32 – Animal evidence showing strong effect of broccoli sprout extract on bladder tumor development in rats.
00:06:06 – Effect of direct supplementation of sulforaphane in prostate cancer patients.
00:07:09 – Bioaccumulation of isothiocyanate metabolites in actual breast tissue.
00:08:32 – Inhibition of breast cancer stem cells.
00:08:53 – History lesson: brassicas were established as having health properties even in ancient Rome.
00:09:16 – Sulforaphane’s ability to enhance carcinogen excretion (benzene, acrolein).
00:09:51 – NRF2 as a genetic switch via antioxidant response elements.
00:10:10 – How NRF2 activation enhances carcinogen excretion via glutathione-S-conjugates.
00:10:34 – Brussels sprouts increase glutathione-S-transferase and reduce DNA damage.
00:11:20 – Broccoli sprout drink increases benzene excretion by 61%.
00:13:31 – Broccoli sprout homogenate increases antioxidant enzymes in the upper airway.
00:15:45 – Cruciferous vegetable consumption and heart disease mortality.
00:16:55 – Broccoli sprout powder improves blood lipids and overall heart disease risk in type 2 diabetics.
00:19:04 – Beginning of aging section.
00:19:21 – Sulforaphane-enriched diet enhances lifespan of beetles from 15 to 30% (in certain conditions).
00:20:34 – Importance of low inflammation for longevity.
00:22:05 – Cruciferous vegetables and broccoli sprout powder seem to reduce a wide variety of inflammatory markers in humans.
00:36:32 – Sulforaphane improves learning in model of type II diabetes in mice.
00:37:19 – Sulforaphane and duchenne muscular dystrophy.
00:37:44 – Myostatin inhibition in muscle satellite cells (in vitro).
00:38:06 – Late-video recap: mortality and cancer, DNA damage, oxidative stress and inflammation, benzene excretion, cardiovascular disease, type II diabetes, effects on the brain (depression, autism, schizophrenia, neurodegeneration), NRF2 pathway.
00:40:27 – Thoughts on figuring out a dose of broccoli sprouts or sulforaphane.
00:41:01 – Anecdotes on sprouting at home.
00:43:14 – On cooking temperatures and sulforaphane activity.
00:43:45 – Gut bacteria conversion of sulforaphane from glucoraphanin.
00:44:24 – Supplements work better when combined with active myrosinase from vegetables.
00:44:56 – Cooking techniques and cruciferous vegetables.
Heating Decreases Epithiospecifier Protein Activity and Increases Sulforaphane Formation in Broccoli
Abstract
Sulforaphane, an isothiocyanate from broccoli, is one of the most potent food-derived anticarcinogens. This compound is not present in the intact vegetable, rather it is formed from its glucosinolate precursor, glucoraphanin, by the action of myrosinase, a thioglucosidase enzyme, when broccoli tissue is crushed or chewed. However, a number of studies have demonstrated that sulforaphane yield from glucoraphanin is low, and that a non-bioactive nitrile analog, sulforaphane nitrile, is the primary hydrolysis product when plant tissue is crushed at room temperature. Recent evidence suggests that in Arabidopsis, nitrile formation from glucosinolates is controlled by a heat-sensitive protein, epithiospecifier protein (ESP), a non-catalytic cofactor of myrosinase. Our objectives were to examine the effects of heating broccoli florets and sprouts on sulforaphane and sulforaphane nitrile formation, to determine if broccoli contains ESP activity, then to correlate heat-dependent changes in ESP activity, sulforaphane content and bioactivity, as measured by induction of the phase II detoxification enzyme quinone reductase (QR) in cell culture. Heating fresh broccoli florets or broccoli sprouts to 60 �C prior to homogenization simultaneously increased sulforaphane formation and decreased sulforaphane nitrile formation. A significant loss of ESP activity paralleled the decrease in sulforaphane nitrile formation. Heating to 70 �C and above decreased the formation of both products in broccoli florets, but not in broccoli sprouts. The induction of QR in cultured mouse hepatoma Hepa lclc7 cells paralleled increases in sulforaphane formation.
Pre-heating broccoli florets and sprouts to 60 �C significantly increased the myrosinase-catalyzed formation of sulforaphane (SF) in vegetable tissue extracts after crushing. This was associated with decreases in sulforaphane nitrile (SF Nitrile) formation and epithiospecifier protein (ESP) activity.
In conclusion, sulforaphane is a phytochemical found in broccoli,and other cruciferous vegetables. An uncontrolled amount of oxidants caused by both internal and external factors can cause oxidative stress in the human body which may ultimately lead to a variety of health issues. Sulforaphane can activate the production of Nrf2, a transcription factor that helps regulate�protective antioxidant mechanisms that control the cell’s response to oxidants. The scope of our information is limited to chiropractic and spinal health issues. To discuss the subject matter, please feel free to ask Dr. Jimenez or contact us at�915-850-0900�.
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Because of this, injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief.
Oxidants are generally produced in a controlled manner in order to regulate essential processes in the human body, including cell division, inflammation, immune function, autophagy, and stress response. However, the uncontrolled production of these oxidants can contribute to oxidative stress, which may affect cellular function, leading to the development of toxicity, chronic disease and cancer. The human body’s protective antioxidant mechanisms are regulated by a series of vital pathways that control the cell’s response to oxidants. The nuclear factor erythroid 2-related factor, otherwise known as Nrf2, is an emerging regulator of cellular resistance to oxidants. The purpose of the article below is to discuss and demonstrate the emerging role of Nrf2 in mitochondrial function.
Abstract
The transcription factor NF-E2 p45-related factor 2 (Nrf2; gene name NFE2L2) allows adaptation and survival under conditions of stress by regulating the gene expression of diverse networks of cytoprotective proteins, including antioxidant, anti-inflammatory, and detoxification enzymes as well as proteins that assist in the repair or removal of damaged macromolecules. Nrf2 has a crucial role in the maintenance of cellular redox homeostasis by regulating the biosynthesis, utilization, and regeneration of glutathione, thioredoxin, and NADPH and by controlling the production of reactive oxygen species by mitochondria and NADPH oxidase. Under homeostatic conditions, Nrf2 affects the mitochondrial membrane potential, fatty acid oxidation, availability of substrates (NADH and FADH2/succinate) for respiration, and ATP synthesis. Under conditions of stress or growth factor stimulation, activation of Nrf2 counteracts the increased reactive oxygen species production in mitochondria via transcriptional upregulation of uncoupling protein 3 and influences mitochondrial biogenesis by maintaining the levels of nuclear respiratory factor 1 and peroxisome proliferator-activated receptor ? coactivator 1?, as well as by promoting purine nucleotide biosynthesis. Pharmacological Nrf2 activators, such as the naturally occurring isothiocyanate sulforaphane, inhibit oxidant-mediated opening of the mitochondrial permeability transition pore and mitochondrial swelling. Curiously, a synthetic 1,4-diphenyl-1,2,3-triazole compound, originally designed as an Nrf2 activator, was found to promote mitophagy, thereby contributing to the overall mitochondrial homeostasis. Thus, Nrf2 is a prominent player in supporting the structural and functional integrity of the mitochondria, and this role is particularly crucial under conditions of stress.
Nrf2 supports the structural and functional integrity of the mitochondria.
Nrf2 activators have beneficial effects when mitochondrial function is compromised.
Introduction
The transcription factor NF-E2 p45-related factor 2 (Nrf2; gene name NFE2L2) regulates the expression of networks of genes encoding proteins with diverse cytoprotective activities. Nrf2 itself is controlled primarily at the level of protein stability. Under basal conditions, Nrf2 is a short-lived protein that is subjected to continuous ubiquitination and proteasomal degradation. There are three known ubiquitin ligase systems that contribute to the degradation of Nrf2. Historically, the first negative regulator of Nrf2 to be discovered was Kelch-like ECH-associated protein 1 (Keap1) [1], a substrate adaptor protein for Cullin 3 (Cul3)/Rbx1 ubiquitin ligase [2], [3], [4]. Keap1 uses a highly efficient cyclic mechanism to target Nrf2 for ubiquitination and proteasomal degradation, during which Keap1 is continuously regenerated, allowing the cycle to proceed (Fig. 1A) [5]. Nrf2 is also subjected to degradation mediated by glycogen synthase kinase (GSK)3/?-TrCP-dependent Cul1-based ubiquitin ligase [6], [7]. Most recently, it was reported that, during conditions of endoplasmic reticulum stress, Nrf2 is ubiquitinated and degraded in a process mediated by the E3 ubiquitin ligase Hrd1 [8].
Figure 1 The cyclic sequential binding and regeneration model for Keap1-mediated degradation of Nrf2. (A) Nrf2 binds sequentially to a free Keap1 dimer: first through its high-affinity ETGE (red sticks) binding domain and then through its low-affinity DLG (black sticks) binding domain. In this conformation of the protein complex, Nrf2 undergoes ubiquitination and is targeted for proteasomal degradation. Free Keap1 is regenerated and able to bind to newly translated Nrf2, and the cycle begins again.(B) Inducers (white diamonds) react with sensor cysteines of Keap1 (blue sticks), leading to a conformational change and impaired substrate adaptor activity. Free Keap1 is not regenerated, and the newly synthesized Nrf2 accumulates and translocates to the nucleus.
In addition to serving as a ubiquitin ligase substrate adaptor protein, Keap1 is also the sensor for a wide array of small-molecule activators of Nrf2 (termed inducers) [9]. Inducers block the cycle of Keap1-mediated degradation of Nrf2 by chemically modifying specific cysteine residues within Keap1 [10], [11] or by directly disrupting the Keap1:Nrf2 binding interface [12], [13]. Consequently, Nrf2 is not degraded, and the transcription factor accumulates and translocates to the nucleus (Fig. 1B), where it forms a heterodimer with a small Maf protein; binds to antioxidant-response elements, the upstream regulatory regions of its target genes; and initiates transcription [14], [15], [16]. The battery of Nrf2 targets comprises proteins with diverse cytoprotective functions, including enzymes of xenobiotic metabolism, proteins with antioxidant and anti-inflammatory functions, and proteasomal subunits, as well as proteins that regulate cellular redox homeostasis and participate in intermediary metabolism.
Nrf2: a Master Regulator of Cellular Redox Homeostasis
The function of Nrf2 as a master regulator of cellular redox homeostasis is widely recognized. The gene expression of both the catalytic and the regulatory subunits of ?-glutamyl cysteine ligase, the enzyme catalyzing the rate-limiting step in the biosynthesis of reduced glutathione (GSH), is directly regulated by Nrf2 [17]. The xCT subunit of system xc-, which imports cystine into cells, is also a direct transcriptional target of Nrf2 [18]. In the cell, cystine undergoes conversion to cysteine, a precursor for the biosynthesis of GSH. In addition to its role in GSH biosynthesis, Nrf2 provides the means for the maintenance of glutathione in its reduced state by the coordinated transcriptional regulation of glutathione reductase 1 [19], [20], which reduces oxidized glutathione to GSH using reducing equivalents from NADPH. The required NADPH is provided by four principal NADPH-generating enzymes, malic enzyme 1 (ME1), isocitrate dehydrogenase 1 (IDH1), glucose-6-phosphate dehydrogenase (G6PD), and 6-phosphogluconate dehydrogenase (PGD), all of which are transcriptionally regulated in part by Nrf2 (Fig. 2) [21], [22], [23], [24]. Curiously, Nrf2 also regulates the inducible gene expression of the cytosolic, microsomal, and mitochondrial forms of aldehyde dehydrogenase [25], which use NAD(P)+ as a cofactor, giving rise to NAD(P)H. Indeed, the levels of NADPH and the NADPH/NADP+ ratio are lower in embryonic fibroblasts isolated from Nrf2-knockout (Nrf2-KO) mice compared to cells from their wild-type (WT) counterparts, and the NADPH levels decrease upon Nrf2 knockdown in cancer cell lines with constitutively active Nrf2 [26]. As expected, the levels of GSH are lower in cells in which Nrf2 has been disrupted; conversely, Nrf2 activation by genetic or pharmacological means leads to GSH upregulation [27], [28], [29]. Importantly, Nrf2 also regulates the gene expression of thioredoxin [30], [31], [32], thioredoxin reductase 1 [28], [29], [32], [33], and sulfiredoxin [34], which are essential for the reduction of oxidized protein thiols.
Figure 2 The role of Nrf2 in the metabolism of rapidly proliferating cells. Nrf2 is a positive regulator of genes encoding enzymes in both the oxidative arm [i.e., glucose-6-phosphate dehydrogenase (G6PD) and 6-phosphogluconate dehydrogenase (PGD)] and the nonoxidative arm [i.e., transaldolase 1 (TALDO1) and transketolase (TKT)] of the pentose phosphate pathway. G6PD and PGD generate NADPH. Nrf2 also regulates the gene expression of the other two NADPH-generating enzymes, malic enzyme 1 (ME1) and isocitrate dehydrogenase 1 (IDH1). The gene expression of phosphoribosyl pyrophosphate amidotransferase (PPAT), which catalyzes the entry into the de novo purine biosynthetic pathway, is also positively regulated by Nrf2, as is the expression of methylenetetrahydrofolate dehydrogenase 2 (MTHFD2), a mitochondrial enzyme with a critical role in providing one-carbon units for de novo purine biosynthesis. Pyruvate kinase (PK) is negatively regulated by Nrf2 and is expected to favor the buildup of glycolytic intermediates and, together with G6PD, metabolite channeling through the pentose phosphate pathway and the synthesis of nucleic acids, amino acids, and phospholipids. Nrf2 negatively regulates the gene expression of ATP-citrate lyase (CL), which may increase the availability of citrate for mitochondrial utilization or (through isocitrate) for IDH1. Red and blue indicate positive and negative regulation, respectively. The mitochondrion is shown in gray. Metabolite abbreviations: G-6-P, glucose 6-phosphate; F-6-P, fructose 6-phosphate; F-1,6-BP, fructose 1,6-bisphosphate; GA-3-P, glyceraldehyde 3-phosphate; 3-PG, 3-phosphoglycerate; PEP, phosphoenolpyruvate; 6-P-Gl, 6-phosphogluconolactone; 6-PG, 6-phosphogluconate; R-5-P, ribulose 5-phosphate; PRPP, 5-phosphoribosyl-?-1-pyrophosphate; THF, tetrahydrofolate; IMP, inosine monophosphate; AMP, adenosine monophosphate; GMP, guanosine monophosphate.
Given the crucial role of Nrf2 as a master regulator of cellular redox homeostasis, it is not surprising that, compared to WT cells, the levels of reactive oxygen species (ROS) are higher in cells in which Nrf2 has been disrupted (Nrf2-KO) [35]. This difference is particularly striking upon challenge with agents causing oxidative stress. Moreover, cells deficient in Nrf2 are much more sensitive to the toxicity of oxidants of various types and cannot be protected by Nrf2 inducers, which, under the same conditions, provide efficient and long-lasting protection to WT cells [29], [36], [37]. In addition to the overall cellular redox homeostasis, Nrf2 is also critical for the maintenance of the mitochondrial redox homeostasis. Thus, compared to WT, the total mitochondrial NADH pool is significantly increased in Keap1-KO and dramatically decreased in Nrf2-KO cells [35].
Using live cell imaging, we recently monitored the rates of ROS production in primary glioneuronal cocultures and brain tissue slices isolated from WT, Nrf2-KO, or Keap1-knockdown (Keap1-KD) mice [38]. As expected, the rate of ROS production was faster in Nrf2-KO cells and tissues compared to their WT counterparts. However, we made the unexpected observation that, compared to WT, Keap1-KD cells also have higher rates of ROS production, although the magnitude of the difference between the WT and the Keap1-KD genotypes was smaller than that between WT and Nrf2-KO. We then analyzed the mRNA levels of NOX2 and NOX4, the catalytic subunits of the two NADPH oxidase (NOX) isoforms that have been implicated in brain pathology, and found that NOX2 is dramatically increased under conditions of Nrf2 deficiency, whereas NOX4 is upregulated when Nrf2 is constitutively activated, although to a smaller extent. Quantitatively, the magnitude of upregulation in cells and tissues from the mutant mice parallels the corresponding increases in ROS production [38]. Interestingly, not only does Nrf2 regulate NADPH oxidase, but the ROS produced by NADPH oxidase can activate Nrf2, as shown in pulmonary epithelial cells and cardiomyocytes [39], [40]. Furthermore, a very recent study has demonstrated that the NADPH oxidase-dependent activation of Nrf2 constitutes an important endogenous mechanism for protection against mitochondrial damage and cell death in the heart during chronic pressure overload [41].
In addition to the catalytic activity of NADPH oxidase, mitochondrial respiration is another major intracellular source of ROS.By use of the mitochondria-specific probe MitoSOX, we have examined the contribution of ROS of mitochondrial origin to the overall ROS production in primary glioneuronal cocultures isolated from WT, Nrf2-KO, or Keap1-KD mice [38]. As expected, Nrf2-KO cells had higher rates of mitochondrial ROS production than WT. In agreement with the findings for the overall ROS production, the rates of mitochondrial ROS production in Keap1-KD were also higher compared to WT cells. Importantly, blocking complex I with rotenone caused a dramatic increase in mitochondrial ROS production in both WT and Keap1-KD cells, but had no effect in Nrf2-KO cells. In contrast to the expected increase in mitochondrial ROS production in WT cells after addition of pyruvate (to enhance the availability of NADH, increase the mitochondrial membrane potential,and normalize respiration), the production of ROS decreased in Nrf2-KO cells. Together, these findings strongly suggest that, in the absence of Nrf2: (i) the activity of complex I is impaired, (ii) the impaired activity of complex I is due to limitation of substrates, and (iii) the impaired activity of complex I is one of the main reasons for the increased mitochondrial ROS production, possibly owing to reverse electron flow from complex II.
Nrf2 Affects Mitochondrial Membrane Potential and Respiration
The mitochondrial membrane potential (??m) is a universal indicator of mitochondrial health and the metabolic state of the cell. In a healthy cell, ??m is maintained by the mitochondrial respiratory chain. Interestingly, a stable isotopic labeling with amino acids in culture-based proteomics study in the estrogen receptor-negative nontumorigenic human breast epithelial MCF10A cell line has shown that the mitochondrial electron transport chain component NDUFA4 is upregulated by pharmacological activation (by sulforaphane) of Nrf2, whereas genetic upregulation of Nrf2 (by Keap1 knockdown) leads to downregulation of the cytochrome c oxidase subunits COX2 and COX4I1 [42]. A study of the liver proteome using two-dimensional gel electrophoresis and matrix-assisted laser desorption/ionization mass spectrometry has found that Nrf2 regulates the expression of ATP synthase subunit ? [43]. In addition, the mitochondrial protein DJ-1, which plays a role in the maintenance of the activity of complex I [44], has been reported to stabilize Nrf2 [45], [46], although the neuroprotective effects of pharmacological or genetic activation of Nrf2 are independent of DJ-1 [47]. However, the consequences of these observations for mitochondrial function have not been investigated.
In agreement with the impaired activity of complex I under conditions of Nrf2 deficiency, the basal ??m is lower in Nrf2-KO mouse embryonic fibroblasts (MEFs) and cultured primary glioneuronal cells in comparison with their WT counterparts (Fig. 3,inset) [35]. In contrast, the basal ??m is higher when Nrf2 is genetically constitutively upregulated (by knockdown or knockout of Keap1). These differences in ??m among the genotypes indicate that respiration is affected by the activity of Nrf2. Indeed, evaluation of the oxygen consumption in the basal state has revealed that, compared to WT, the oxygen consumption is lower in Nrf2-KO and Keap1-KO MEFs, by ~50 and ~35%, respectively.
Figure 3 Proposed mechanism for compromised mitochondrial function under conditions of Nrf2 deficiency. (1) The decreased levels of ME1, IDH1, G6PD, and PGD result in lower NADPH levels. (2) The levels of GSH are also low. (3) The low activity of ME1 may decrease the pool of pyruvate entering the mitochondria. (4) The generation of NADH is slower, leading to impaired activity of complex I and increased mitochondrial ROS production. (5) The reduction of FAD to FADH2 in mitochondrial proteins is also decreased, lowering the electron flow from FADH2 to UbQ and into complex III. (6) The slower formation of UbQH2 may lower the enzyme activity of succinate dehydrogenase. (7) The increased levels of ROS may further inhibit the activity of complex II. (8) The lower efficiency of fatty acid oxidation contributes to the decreased substrate availability for mitochondrial respiration. (9) Glycolysis is enhanced as a compensatory mechanism for the decreased ATP production in oxidative phosphorylation. (10) ATP synthase operates in reverse to maintain ??m. Red and blue indicate upregulation and downregulation, respectively. The boxes signify availability of experimental evidence. The inset shows images of mitochondria of WT and Nrf2-KO cortical astrocytes visualized by the potentiometric fluorescent probe tetramethylrhodamine methyl ester (TMRM; 25 nM). Scale bar, 20 �m.
These differences in ??m and respiration among the genotypes are reflected by the rate of utilization of substrates for mitochondrial respiration. Application of substrates for the tricarboxylic acid (TCA) cycle (malate/pyruvate, which in turn increase the production of the complex I substrate NADH) or methyl succinate, a substrate for complex II, causes a stepwise increase in ??m in both WT and Keap1-KD neurons, but the rate of increase is higher in Keap1-KD cells. More importantly, the shapes of the response to these TCA cycle substrates are different between the two genotypes, whereby the rapid rise in ??m in Keap1-KD cells upon substrate addition is followed by a quick drop rather than a plateau, suggesting an unusually fast substrate consumption. These findings are in close agreement with the much lower (by 50�70%) levels of malate, pyruvate, and succinate that have been observed after a 1-h pulse of [U-13C6]glucose in Keap1-KO compared to WT MEF cells [24]. In Nrf2-KO neurons, only pyruvate is able to increase the ??m, whereas malate and methyl succinate cause mild depolarization. The effect of Nrf2 on mitochondrial substrate production seems to be the main mechanism by which Nrf2 affects mitochondrial function. The mitochondrial NADH redox index (the balance between consumption of NADH by complex I and production of NADPH in the TCA cycle) is significantly lower in Nrf2-KO cells in comparison with their WT counterparts, and furthermore, the rates of regeneration of the pools of NADH and FADH2 after inhibition of complex IV (by use of NaCN) are slower in the mutant cells.
In mitochondria isolated from murine brain and liver, supplementation of substrates for complex I or for complex II increases the rate of oxygen consumption more strongly when Nrf2 is activated and less efficiently when Nrf2 is disrupted [35]. Thus, malate induces a higher rate of oxygen consumption in Keap1-KD compared to WT, but its effect is weaker in Nrf2-KO mitochondria. Similarly, in the presence of rotenone (when complex I is inhibited), succinate activates oxygen consumption to a greater extent in Keap1-KD compared to WT, whereas the response in Nrf2-KO mitochondria is diminished. In addition, Nrf2-KO primary neuronal cultures and mice are more sensitive to the toxicity of the complex II inhibitors 3-nitropropionic acid and malonate, whereas intrastriatal transplantation of Nrf2-overexpressing astrocytes is protective [48], [49]. Similarly, Nrf2-KO mice are more sensitive to, whereas genetic or pharmacological activation of Nrf2 has protective effects against, neurotoxicity caused by the complex I inhibitor 1-methyl-4-phenylpyridinium ion in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine animal model of Parkinson?s disease [49], [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61].
The respiratory control ratio (RCR), the ratio of State 3 (ADP-stimulated) to State 4 respiration (no ADP present), is decreased in the absence of Nrf2, but the RCR is similar between Keap1-KD and WT mitochondria [35]. As the RCR is an indication of the degree of coupling of the mitochondrial respiratory chain activity to oxidative phosphorylation, this finding indicates that the higher rate of respiration in Keap1-KD mitochondria is not due to uncoupling of oxidative phosphorylation. It further suggests that oxidative phosphorylation is more efficient when Nrf2 is activated. The higher rate of respiration in Keap1-KD mitochondria is consistent with the higher levels of mitochondrial ROS production [38] as higher respiration rates may lead to increased electron leak. However, under conditions of oxidative stress, the increased ROS production is counteracted by the Nrf2-dependent transcriptional upregulation of uncoupling protein 3 (UCP3), which increases the proton conductance of the mitochondrial inner membrane and consequently decreases the production of superoxide [62]. Very recently, it was shown that the lipid peroxidation product 4-hydroxy-2-nonenal mediates the Nrf2-dependent upregulation of UCP3 in cardiomyocytes; this might be particularly important for protection under conditions of oxidative stress such as those during ischemia�reperfusion [63].
Nrf2 Affects the Efficiency of Oxidative Phosphorylation and the Synthesis of ATP
In agreement with the effect of Nrf2 on respiration, in brain and liver mitochondria, Nrf2 deficiency results in a decreased efficiency of oxidative phosphorylation (as estimated by the ratio of ADP to oxygen, which is consumed for ATP synthesis), whereas Nrf2 activation (Keap1-KD) has the opposite effect [35]. Compared to WT, the ATP levels are significantly higher in cells with constitutive upregulation of Nrf2 and lower when Nrf2 is knocked down [64] or disrupted [35]. Furthermore, the use of inhibitors of oxidative phosphorylation (oligomycin) or glycolysis (iodoacetic acid) has revealed that Nrf2 changes the way by which cells produce ATP. Thus, in WT neurons, oligomycin causes a complete drop in ATP and iodoacetic acid has no further effect. Remarkably, in Nrf2-KO cells, oligomycin increases the ATP levels, which are then slowly, but completely, depleted by iodoacetic acid, indicating that in the absence of Nrf2, glycolysis, and not oxidative phosphorylation, is the main source of ATP production. Interestingly, despite the increased efficiency of oxidative phosphorylation in Keap1-KD cells, addition of oligomycin results in an ~80% decrease in ATP levels, and iodoacetic acid causes a further ~20% decrease. Thus, either Nrf2 deficiency or its constitutive activation reduces the contribution of oxidative phosphorylation and increases the contribution of glycolysis toward the synthesis of ATP. This effect is particularly pronounced when Nrf2 is absent and is consistent with the dependence of the ??m on the presence of glucose in the medium [35] and the increased levels of glycolytic intermediates (G-6-P, F-6-P, dihydroxyacetone phosphate, pyruvate, and lactate) after knockdown of Nrf2 [24].
The increase in ATP levels after inhibition of the F1F0-ATPase by oligomycin indicates that in the absence of Nrf2, the F1F0-ATPase functions as an ATPase and not an ATP synthase, i.e., it operates in reverse. Such reversal in activity most likely reflects the need to pump protons across the inner mitochondrial membrane in an attempt to maintain the ??m, which is crucial for the functional integrity of this organelle. The reversal of the function of the F1F0-ATPase is also evidenced by the observed mitochondrial depolarization upon oligomycin administration to Nrf2-KO cells, which is in sharp contrast to the hyperpolarization occurring in their WT or Keap1-deficient counterparts [35]. Overall, it seems that under conditions of Nrf2 deficiency ATP is produced primarily in glycolysis, and this ATP is then used in part by the F1F0-ATPase to maintain the ??m.
Nrf2 Enhances Mitochondrial Fatty Acid Oxidation
The effect of Nrf2 deficiency on the ??m is particularly pronounced when cells are incubated in medium without glucose, and the ??m is ~50% lower in Nrf2-KO compared to WT cells [35]. Under conditions of glucose deprivation, mitochondrial fatty acid oxidation (FAO) is a major provider of substrates for respiration and oxidative phosphorylation, suggesting that Nrf2 may affect FAO. Indeed, the efficiency of FAO for both the long-chain (C16:0) saturated fatty acid palmitic acid and the short-chain (C6:0) hexanoic acid is higher in Keap1-KO MEFs and isolated heart and liver mitochondria than in their WT counterparts, whereas it is lower in Nrf2-KO cells and mitochondria [65]. These effects are also highly relevant to humans: indeed, metabolic changes indicative of better integration of FAO with the activity of the TCA cycle have been reported to occur in human intervention studies with diets rich in glucoraphanin, the precursor of the classical Nrf2 activator sulforaphane [66].
During the first step of mitochondrial FAO, the pro-R hydrogen of the ?-carbon leaves as a hydride that reduces the FAD cofactor to FADH2, which in turn transfers electrons to ubiquinone (UbQ) in the respiratory chain, ultimately contributing to ATP production. Whereas stimulation of FAO by palmitoylcarnitine in the absence of glucose causes the expected increase in the ATP levels in WT and Keap1-KO cells, with the ATP rise being faster in Keap1-KO cells, the identical treatment produces no ATP changes in Nrf2-KO MEFs [65]. This experiment demonstrates that, in the absence of Nrf2, FAO is suppressed, and furthermore, it implicates suppression of FAO as one of the reasons for the lower ATP levels under conditions of Nrf2 deficiency [35], [64].
Notably, human 293 T cells in which Nrf2 has been silenced have a lower expression of CPT1 and CPT2[67], two isoforms of carnitine palmitoyltransferase (CPT), the rate-limiting enzyme in mitochondrial FAO. In agreement, the mRNA levels of Cpt1 are lower in livers of Nrf2-KO compared to WT mice [68]. CPT catalyzes the transfer of the acyl group of a long-chain fatty acyl-CoA from coenzyme A to l-carnitine and thus permits the import of acylcarnitine from the cytoplasm into the mitochondria. Although this has not been examined to date, it is possible that in addition to the transcriptional effects on CPT1 expression, Nrf2 may also affect the function of this enzyme by controlling the levels of its main allosteric inhibitor, malonyl-CoA. This is because, by a mechanism that is currently unclear, Nrf2 regulates negatively the expression of stearoyl CoA desaturase (SCD) [69] and citrate lyase (CL) [69], [70]. Curiously, knockout or inhibition of SCD leads to increased phosphorylation and activation of AMP-activated protein kinase (AMPK) [71], [72], [73], and it can be speculated that, in the absence of Nrf2, the SCD levels will increase, in turn lowering AMPK activity. This could be further compounded by the reduced protein levels of AMPK that have been observed in livers of Nrf2-KO mice [68], a finding that is in close agreement with the increased AMPK levels, which have been reported in livers of Keap1-KD mice [74]. One consequence of the decreased AMPK activity is the relief of its inhibitory phosphorylation (at Ser79) of acetyl-CoA carboxylase (ACC) [75], which could be further transcriptionally upregulated in the absence of Nrf2 because it is downregulated by Nrf2 activation [70]. The high ACC activity, in combination with the upregulated CL expression that will increase the production of acetyl-CoA, the substrate for ACC, may ultimately increase the levels of the ACC product, malonyl-CoA. The high levels of malonyl-CoA will inhibit CPT, thereby decreasing the transport of fatty acids into the mitochondria. Finally, Nrf2 positively regulates the expression of CD36 [76], a translocase that imports fatty acids across plasma and mitochondrial membranes. Thus, one mechanism by which Nrf2 may affect the efficiency of mitochondrial FAO is by regulating the import of long-chain fatty acids into the mitochondria.
In addition to direct transcriptional regulation, Nrf2 may also alter the efficiency of mitochondrial FAO by its effects on the cellular redox metabolism. This may be especially relevant when Nrf2 activity is low or absent, conditions that shift the cellular redox status toward the oxidized state. Indeed, several FAO enzymes have been identified as being sensitive to redox changes. One such enzyme is very long-chain acyl-CoA dehydrogenase (VLCAD), which contributes more than 80% to the palmitoyl-CoA dehydrogenation activity in human tissues [77]. Interestingly, Hurd et al. [78] have shown that VLCAD contains cysteine residues that significantly change their redox state upon exposure of isolated rat heart mitochondria to H2O2. Additionally, S-nitrosylation of murine hepatic VLCAD at Cys238 improves the catalytic efficiency of the enzyme [79], and it is likely that oxidation of the same cysteine may have the opposite effect, ultimately lowering the efficiency of mitochondrial FAO. It is therefore possible that, although the expression levels of VLCAD are not significantly different in WT, Nrf2-KO, or Keap1-KO MEFs [65], the enzyme activity of VLCAD could be lower in the absence of Nrf2 owing to the higher levels of ROS.
Based on all of these findings, it can be proposed that (Fig. 3): in the absence of Nrf2, the NADPH levels are lower owing to decreased expression of ME1, IDH1, G6PD, and PGD. The levels of reduced glutathione are also lower owing to decreased expression of enzymes that participate in its biosynthesis and regeneration and the lower levels of NADPH that are required for the conversion of the oxidized to the reduced form of glutathione. The low expression of ME1 will decrease the pool of pyruvate entering the mitochondria, with glycolysis becoming the major source of pyruvate. The generation of NADH is slower, leading to impaired activity of complex I and increased mitochondrial ROS production. The reduction of FAD to FADH2 is also slower, at least in part owing to less efficient fatty acid oxidation, compromising the electron flow from FADH2 to UbQ and into complex III. As UbQH2 is an activator of succinate dehydrogenase [80], slowing down its formation may lower the enzyme activity of succinate dehydrogenase. The increased levels of superoxide and hydrogen peroxide can inhibit complex II activity further [81]. The lower efficiency of fatty acid oxidation contributes to the decreased substrate availability for mitochondrial respiration and ATP production in oxidative phosphorylation. As a compensatory mechanism, glycolysis is enhanced. ATP synthase functions in reverse, as an ATPase, in an attempt to maintain the ??m.
Nrf2 and Mitochondrial Biogenesis
It has been reported that, compared to WT, the livers of Nrf2-KO mice have a lower mitochondrial content (as determined by the ratio of mitochondrial to nuclear DNA); this is further decreased by a 24-h fast in both WT and Nrf2-KO mice; in contrast, although no different from WT under normal feeding conditions, the mitochondrial content in mice with high Nrf2 activity is not affected by fasting [82]. Interestingly, supplementation with the Nrf2 activator (R)-?-lipoic acid [83], [84], [85] promotes mitochondrial biogenesis in 3T3-L1 adipocytes [86]. Two classes of nuclear transcriptional regulators play critical roles in mitochondrial biogenesis. The first class are transcription factors, such as nuclear respiratory factors11 and 2, which control the expression of genes encoding subunits of the five respiratory complexes, mitochondrial translational components, and heme biosynthetic enzymes that are localized to the mitochondrial matrix [88]. Piantadosi et al. [89] have shown that the Nrf2-dependent transcriptional upregulation of nuclear respiratory factor 1 promotes mitochondrial biogenesis and protects against the cytotoxicity of the cardiotoxic anthracycline chemotherapeutic agent doxorubicin. In contrast, Zhang et al. [82] have reported that genetic activation of Nrf2 does not affect the basal mRNA expression of nuclear respiratory factor 1 in the murine liver.
The second class of nuclear transcriptional regulators with critical functions in mitochondrial biogenesis are transcriptional coactivators, such as peroxisome proliferator-activated receptor ? coactivators (PGC)1? and 1?, which interact with transcription factors, the basal transcriptional and RNA-splicing machinery, and histone-modifying enzymes [88], [90], [91]. The expression of the PGC1 family of coactivators is influenced by numerous environmental signals. Treatment of human fibroblasts with the Nrf2 activator sulforaphane causes an increase in mitochondrial mass and induction of PGC1? and PGC1? [92], although the potential dependence on Nrf2 was not examined in this study. However, diabetic mice in which Nrf2 is either activated by Keap1 gene hypomorphic knockdown (db/db:Keap1flox/?:Nrf2+/+) or disrupted (db/db:Keap1flox/?:Nrf2?/?) have lower hepatic PGC1? expression levels than control animals (db/db:Keap1flox/+:Nrf2+/+) [93]. No differences in the mRNA levels for PGC1? are seen in livers of nondiabetic mice that are either WT or Nrf2-KO, whereas these levels are lower in Nrf2-overexpressing (Keap1-KD and liver-specific Keap1-KO) animals [82]. Notably, a 24-h fast increases the levels of PGC1? mRNA in the livers of mice of all genotypes, but the increase is significantly greater in livers of Nrf2-KO compared to WT or Nrf2-overexpressing mice. Compared to WT, Nrf2-KO mice experiencing septic infection or acute lung injury due to infection show attenuated transcriptional upregulation of nuclear respiratory factor 1 and PGC1? [94], [95]. Together, these observations suggest that the role of Nrf2 in maintaining the levels of both nuclear respiratory factor 1 and PGC1? is complex and becomes most prominent under conditions of stress.
In addition to expression of genes encoding mitochondrial proteins, mitochondrial biogenesis requires the synthesis of nucleotides. Genetic activation of Nrf2 enhances purine biosynthesis by upregulating the pentose phosphate pathway and the metabolism of folate and glutamine, particularly in rapidly proliferating cells (Fig. 2) [24]. Analysis of the transcriptome of mutant Drosophila deficient for the mitochondrial serine/threonine protein kinase PTEN-induced putative kinase 1 (PINK1) has shown that mitochondrial dysfunction leads to the transcriptional upregulation of genes affecting nucleotide metabolism [96], suggesting that the enhanced nucleotide biosynthesis represents a mechanism for protection against the neurotoxic consequences of PINK1 deficiency. Nrf2 regulates the expression of phosphoribosyl pyrophosphate amidotransferase (PPAT), which catalyzes the entry into the de novo purine nucleotide biosynthetic pathway, and mitochondrial methylenetetrahydrofolate dehydrogenase 2 (MTHFD2) (Fig. 2). The latter is a bifunctional enzyme with dehydrogenase and cyclohydrolase activities that is critical in providing both glycine and formate as sources of one-carbon units for purine biosynthesis in rapidly growing cells [97]. It is therefore likely that Nrf2 activation might be protective and might reverse mitochondrial dysfunction in PINK1 deficiency. Indeed, pharmacological activation of Nrf2 by sulforaphane, or the triterpenoid RTA-408, restores ??m and protects PINK1-deficient cells against dopamine toxicity [98]. Although the underlying mechanisms seem to be complex, together, these findings indicate that Nrf2 activity may affect mitochondrial biogenesis by influencing the expression levels of critical transcription factors and coactivators, as well as by enhancing nucleotide biosynthesis.
Nrf2 and Mitochondrial Integrity
Although direct evidence is not always available, there are strong indications that Nrf2 is important for mitochondrial integrity, particularly under conditions of oxidative stress. Mitochondria isolated from the brain and liver of rats that had been administered a single dose of the Nrf2 activator sulforaphane are resistant to opening of the mitochondrial permeability transition pore (mPTP) caused by the oxidant tert-butylhydroperoxide [99], [100]. The mPTP, a complex that allows the mitochondrial inner membrane to become permeable to molecules with masses up to 1500 Da, was recently identified to be formed from dimers of the F0F1-ATP synthase [101]. The sulforaphane-mediated resistance to mPTP opening correlates with increased antioxidant defenses, and the levels of mitochondrial GSH, glutathione peroxidase 1, malic enzyme 3, and thioredoxin 2 are all upregulated in mitochondrial fractions isolated from sulforaphane-treated animals [100].
Mitochondrial protein damage and impairment in respiration caused by the electrophilic lipid peroxidation product 4-hydroxy-2-nonenal are attenuated in mitochondria isolated from the cerebral cortex of sulforaphane-treated mice [102]. In rat renal epithelial cells and in kidney, sulforaphane is protective against cisplatin- and gentamicin-induced toxicity and loss of ??m[103], [104]. Protection against a panel of oxidants (superoxide, hydrogen peroxide, peroxynitrite) and electrophiles (4-hydroxy-2-nonenal and acrolein) and an increase in mitochondrial antioxidant defenses have been also observed upon treatment of rat aortic smooth muscle cells with sulforaphane [105]. In a model of contrast-induced acute kidney injury, limb ischemic preconditioning was recently shown to have protective effects, including inhibition of the opening of the mPTP and mitochondrial swelling, by activation of Nrf2 consequent to the inhibition of GSK3? [106].
Mitophagy, the process by which dysfunctional mitochondria are selectively engulfed by autophagosomes and delivered to lysosomes to be degraded and recycled by the cell, is essential for mitochondrial homeostasis [107], [108]. Whereas no causative relation between Nrf2 and mitophagy has been established, there is evidence that the transcription factor may be important in mitochondrial quality control by playing a role in mitophagy. This might be especially prominent under conditions of oxidative stress. Thus, in a model of sepsis, the increases in the levels of the autophagosome marker MAP1 light chain 3-II (LC3-II) and the cargo protein p62 at 24 h postinfection are suppressed in Nrf2-KO compared to WT mice [109]. A small-molecule inducer of mitophagy (called p62-mediated mitophagy inducer, PMI) was recently discovered; this 1,4-diphenyl-1,2,3-triazole compound was originally designed as an Nrf2 activator that disrupts the interaction of the transcription factor with Keap1 [110]. Similar to cells in which Nrf2 is genetically upregulated (Keap1-KD or Keap1-KO), cells exposed to PMI have higher resting ??m. Importantly, the increase in mitochondrial LC3 localization that is observed after PMI treatment of WT cells does not occur in Nrf2-KO cells, suggesting the involvement of Nrf2.
Last, ultrastructural analysis of liver sections has revealed the presence of swollen mitochondria with reduced crista and disrupted membranes in hepatocytes of Nrf2-KO, but not WT, mice that had been fed a high-fat diet for 24 weeks; notably, these livers show clear evidence of oxidative stress and inflammation [68]. It can be concluded that Nrf2 has a critical role in maintaining mitochondrial integrity under conditions of oxidative and inflammatory stress.
Sulforaphane and Its Effects on Cancer, Mortality, Aging, Brain and Behavior, Heart Disease & More
Isothiocyanates are some of the most important plant compounds you can get in your diet. In this video I make the most comprehensive case for them that has ever been made. Short attention span? Skip to your favorite topic by clicking one of the time points below. Full timeline below.
Key sections:
00:01:14 – Cancer and mortality
00:19:04 – Aging
00:26:30 – Brain and behavior
00:38:06 – Final recap
00:40:27 – Dose
Full timeline:
00:00:34 – Introduction of sulforaphane, a major focus of the video.
00:01:14 – Cruciferous vegetable consumption and reductions in all-cause mortality.
00:02:12 – Prostate cancer risk.
00:02:23 – Bladder cancer risk.
00:02:34 – Lung cancer in smokers risk.
00:02:48 – Breast cancer risk.
00:03:13 – Hypothetical: what if you already have cancer? (interventional)
00:03:35 – Plausible mechanism driving the cancer and mortality associative data.
00:04:38 – Sulforaphane and cancer.
00:05:32 – Animal evidence showing strong effect of broccoli sprout extract on bladder tumor development in rats.
00:06:06 – Effect of direct supplementation of sulforaphane in prostate cancer patients.
00:07:09 – Bioaccumulation of isothiocyanate metabolites in actual breast tissue.
00:08:32 – Inhibition of breast cancer stem cells.
00:08:53 – History lesson: brassicas were established as having health properties even in ancient Rome.
00:09:16 – Sulforaphane’s ability to enhance carcinogen excretion (benzene, acrolein).
00:09:51 – NRF2 as a genetic switch via antioxidant response elements.
00:10:10 – How NRF2 activation enhances carcinogen excretion via glutathione-S-conjugates.
00:10:34 – Brussels sprouts increase glutathione-S-transferase and reduce DNA damage.
00:11:20 – Broccoli sprout drink increases benzene excretion by 61%.
00:13:31 – Broccoli sprout homogenate increases antioxidant enzymes in the upper airway.
00:15:45 – Cruciferous vegetable consumption and heart disease mortality.
00:16:55 – Broccoli sprout powder improves blood lipids and overall heart disease risk in type 2 diabetics.
00:19:04 – Beginning of aging section.
00:19:21 – Sulforaphane-enriched diet enhances lifespan of beetles from 15 to 30% (in certain conditions).
00:20:34 – Importance of low inflammation for longevity.
00:22:05 – Cruciferous vegetables and broccoli sprout powder seem to reduce a wide variety of inflammatory markers in humans.
00:36:32 – Sulforaphane improves learning in model of type II diabetes in mice.
00:37:19 – Sulforaphane and duchenne muscular dystrophy.
00:37:44 – Myostatin inhibition in muscle satellite cells (in vitro).
00:38:06 – Late-video recap: mortality and cancer, DNA damage, oxidative stress and inflammation, benzene excretion, cardiovascular disease, type II diabetes, effects on the brain (depression, autism, schizophrenia, neurodegeneration), NRF2 pathway.
00:40:27 – Thoughts on figuring out a dose of broccoli sprouts or sulforaphane.
00:41:01 – Anecdotes on sprouting at home.
00:43:14 – On cooking temperatures and sulforaphane activity.
00:43:45 – Gut bacteria conversion of sulforaphane from glucoraphanin.
00:44:24 – Supplements work better when combined with active myrosinase from vegetables.
00:44:56 – Cooking techniques and cruciferous vegetables.
00:46:06 – Isothiocyanates as goitrogens.
Nrf2 is a transcription factor which plays an important role in the cellular antioxidant defense system of the human body. The antioxidant responsive element, or ARE, is a regulatory mechanism of genes. Many research studies have demonstrated that Nrf2, or NF-E2-related factor 2, regulates a wide variety of ARE-driven genes throughout several types of cells. Nrf2 was also found to play an essential role in cellular protection and anti-carcinogenicity, which demonstrates that Nrf2 may be an effective treatment in the management of neurodegenerative diseases and cancers believed to be caused by oxidative stress. Dr. Alex Jimenez D.C., C.C.S.T. Insight
Concluding Remarks
Although many questions still remain open, the available experimental evidence clearly indicates that Nrf2 is an important player in the maintenance of mitochondrial homeostasis and structural integrity. This role becomes particularly critical under conditions of oxidative, electrophilic, and inflammatory stress when the ability to upregulate Nrf2-mediated cytoprotective responses influences the overall health and survival of the cell and the organism. The role of Nrf2 in mitochondrial function represents another layer of the broad cytoprotective mechanisms orchestrated by this transcription factor. As many human pathological conditions have oxidative stress, inflammation, and mitochondrial dysfunction as essential components of their pathogenesis, pharmacological activation of Nrf2 holds promise for disease prevention and treatment. Comprehensive understanding of the precise mechanisms by which Nrf2 affects mitochondrial function is essential for rational design of future clinical trials and may offer new biomarkers for monitoring therapeutic efficacy.
The purpose of the article above was to discuss�as well as demonstrate�the emerging role of Nrf2 in mitochondrial function. Nrf2, or nuclear factor erythroid 2-related factor, is an emerging regulator of cellular resistance to oxidants which can contribute to oxidative stress, affecting cellular function and leading to the development of toxicity, chronic disease, and even cancer. While the production of oxidants in the human body can serve�various purposes,�including cell division, inflammation, immune function, autophagy, and stress response, it’s essential to control their overproduction to prevent health issues. The scope of our information is limited to chiropractic and spinal health issues. To discuss the subject matter, please feel free to ask Dr. Jimenez or contact us at�915-850-0900�.
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Because of this, injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief. �
Neurodegenerative diseases, such as Alzheimer’s disease and Parkinson’s disease, affect millions of individuals worldwide. A variety of treatment options are available to treat the symptoms of several neurodegenerative diseases although the results are often limited. Research studies have found that oxidative stress caused by both internal and external factors can be a cause for the development of neurodegenerative diseases. The transcription factor, Nrf2, has been determined to function as a major defense mechanism against oxidative stress. The purpose of the article below is to show the effects of Nrf2 on neurodegenerative diseases.
Modulation of Proteostasis by Transcription Factor NRF2
Neurodegenerative diseases are linked to the accumulation of specific protein aggregates, suggesting an intimate connection between injured brain and loss of proteostasis. Proteostasis refers to all the processes by which cells control the abundance and folding of the proteome thanks to a wide network that integrates the regulation of signaling pathways, gene expression and protein degradation systems. This review attempts to summarize the most relevant findings about the transcriptional modulation of proteostasis exerted by the transcription factor NRF2 (nuclear factor (erythroid-derived 2)-like 2). NRF2 has been classically considered as the master regulator of the antioxidant cell response, although it is currently emerging as a key component of the transduction machinery to maintain proteostasis. As we will discuss, NRF2 could be envisioned as a hub that compiles emergency signals derived from misfolded protein accumulation in order to build a coordinated and perdurable transcriptional response. This is achieved by functions of NRF2 related to the control of genes involved in the maintenance of the endoplasmic reticulum physiology, the proteasome and autophagy.
Keywords:Neurodegenerative diseases, Unfolded protein response, Proteasome, Ubiquitin, Autophagy, Oxidative stress
Nuclear Factor (erythroid-derived 2)-like 2 (NRF2) is a basic-leucine-zipper protein considered nowadays as a master regulator of cellular homeostasis. It controls the basal and stress-inducible expression of over 250 genes that share in common a cis-acting enhancer termed the antioxidant response element (ARE) [1], [2], [3], [4], [5]. These genes participate in phase I, II and III detoxification reactions, glutathione and peroxiredoxin/thioredoxin metabolism, NADPH production through the pentose phosphate pathway and malic enzyme, fatty acid oxidation, iron metabolism, and proteostasis [6]. Given these wide cytoprotective functions, it is possible that a single pharmacological hit in NRF2 might mitigate the effect of the main culprits of chronic diseases, including oxidative, inflammatory and proteotoxic stress. The role of NRF2 in the modulation of the antioxidant defense and resolution of inflammation have been addressed in numerous studies (reviewed in [7]). Here, we will focus on its role in proteostasis, i.e., the homeostatic control of protein synthesis, folding, trafficking and degradation. Examples will be provided in the context of neurodegenerative diseases.
Loss of Proteostasis Influences NRF2 Activity in Neurodegenerative Diseases
A general hallmark of neurodegenerative diseases is the occurrence of aberrant aggregation of some proteins. Thus, misfolded protein aggregates of ?-synuclein (?-SYN) are found in Parkinson’s disease (PD), ?-amyloid (A?) plaques and hyper-phosphorylated TAU neurofibrillary tangles in Alzheimer’s disease (AD), huntingtin (Htt) in Huntington’s disease (HD), superoxide dismutase 1 (SOD1) and TAR DNA binding protein 43 (TDP-43) in amyotrophic lateral sclerosis (ALS), prion protein (PrP) in spongiform encephalopathies, etc. Protein aggregates can have an impact on several cellular pathways, which in turn may affect NRF2 levels and activity.
Different Layers of Regulation Tightly Control NRF2 Activity
Under physiological conditions, cells exhibit low NRF2 protein levels because of its rapid turnover. In response to different stimuli, NRF2 protein is accumulated, enters the nucleus and increases the transcription of ARE-containing genes. Therefore, management of NRF2 protein levels is a key point that should integrate positive and negative input signals. As we will discuss further, NRF2 is activated by diverse overlapping mechanisms to orchestrate a rapid and efficient response but on the other hand NRF2 could be inhibited, probably in a second phase, in order to switch off its response.
From the classic point of view, activation of NRF2 has been considered as a consequence of the cellular response to oxidant or electrophilic compounds. In this regard, the ubiquitin E3 ligase adaptor Kelch-like ECH-associated protein 1 (KEAP1) plays a crucial role. Molecular details will be further addressed in Section 4.1. In brief, KEAP1 acts as a redox sensor due to critical cysteine residues leading to NRF2 ubiquitination and proteasomal degradation. In addition to this classic modulation, NRF2 is profoundly regulated by signaling events. Indeed, different kinases have been shown to phosphorylate and regulate NRF2. For instance, NRF2 can be phosphorylated by mitogen activated protein kinases (MAPKs), although its contribution to NRF2 activity remains unclear [8], [9], [10], [11]. PKA kinase as well as some PKC isozymes [12], CK2 [13] or Fyn [14] phosphorylate NRF2 modifying its stability. Previous work from our group reported that glycogen synthase kinse-3? (GSK-3?) inhibits NRF2 by nuclear exclusion and proteasomal degradation [15], [25], [26], [27], [28], [29], [30]. The molecular details will be discussed in the Section 4.1. Furthermore, NRF2 is submitted to other types of regulation. For instance, NRF2 acetylation by CBP/p300 increases its activity [17], while it is inhibited by miR153, miR27a, miR142-5p, and miR144 [16], or by methylation of cytosine-guanine (CG) islands within the NRF2 promoter [18].
Impact of Protein Aggregates on NRF2 Regulatory Mechanisms
In this section we will focus in how accumulation of misfolded protein could impact NRF2 activity providing some of the pathways mentioned above as illustrative examples. Firstly, we need to consider that protein accumulation has been tightly linked with oxidative damage. Indeed, misfolded protein accumulation and aggregation induce abnormal production of reactive oxygen species (ROS) from mitochondria and other sources [19]. As mentioned above, ROS will modify redox-sensitive cysteines of KEAP1 leading to the release, stabilization and nuclear localization of NRF2.
Regarding proteinopathies, an example of dysregulated signaling events that may affect NRF2 is provided by the hyperactivation of GSK-3? in AD. GSK-3?, also known as TAU kinase, participates in the phosphorylation of this microtubule-associated protein, resulting in its aggregation, formation of neurofibrillary tangles and interruption of axonal transport (reviewed in [20]). On the other hand, GSK-3? dramatically reduces NRF2 levels and activity as mentioned above. Although not widely accepted, the amyloid cascade proposes that toxic A? oligomers increase GSK-3? activity together with TAU hyper-phosphorylation and neuron death [21], [22]. There are different models to explain how A? favors GSK3-? activity. For instance, A? binds to the insulin receptor and inhibits PI3K and AKT signaling pathways, which are crucial to maintain GSK-3? inactivated by phosphorylation at its N-terminal Ser9 residue [23]. On the other hand, extracellular A? interacts with Frizzled receptors, blocking WNT signaling [24] and again resulting in release of active GSK-3?. In summary, A? accumulation leads to abnormal hyperactivation of GSK-3?, thus impairing an appropriate NRF2 response.
As discussed in the following section, misfolded proteins lead to activation of PERK and MAPKs, which in turn up-regulate NRF2 [31], [8], [9], [10], [11]. Moreover, dysregulated CBP/p300 activity has been reported in several proteinopathies [32] and a global decrease in DNA methylation in AD brains was also shown [33], therefore providing grounds to explore the relevance of these findings in NRF2 regulation.
We and others have observed in necropsies of PD and AD patients an increase in NRF2 protein levels and some of its targets, such as heme oxygenase 1 (HMOX1), NADPH quinone oxidase 1 (NQO1), p62, etc., both by immunoblot and by immunohistochemistry [34], [35], [36], [37], [38], [39]. The up-regulation of NRF2 in these diseases is interpreted as an unsuccessful attempt of the diseased brain to recover homeostatic values. However, another study indicated that NRF2 is predominantly localized in the cytoplasm of AD hippocampal neurons, suggesting reduced NRF2 transcriptional activity in the brain [40]. It is conceivable that the disparity of these observations is related to changes in the factors that control NRF2 along the progressive stages of neurodegeneration.
Three major systems contribute to proteostasis, namely the unfolded protein response (UPR), the ubiquitin proteasome system (UPS) and autophagy. Next, we present evidence to envision NRF2 as a hub connecting emergency signals activated by protein aggregates with the protein derivative machinery.
NRF2 Participates in the Unfolded Protein Response (UPR)
NRF2 Activation in Response to the UPR
Oxidative protein folding in the ER is driven by a number of distinct pathways, the most conserved of which involves the protein disulfide-isomerase (PDI) and the sulfhydryl oxidase endoplasmic oxidoreductin 1 (ERO1? and ERO1? in mammals) as disulfide donor. Briefly, PDI catalyzes the formation and breakage of disulfide bonds between cysteine residues within proteins, as they fold, due to the reduction and oxidation of its own cysteine aminoacids. PDI is recycled by the action of the housekeeping enzyme ERO1, which reintroduces disulfide bonds into PDI [41]. Molecular oxygen is the terminal electron acceptor of ERO1, which generates stoichiometric amounts of hydrogen peroxide for every disulfide bond produced [42]. Peroxidases (PRX4) and glutathione peroxidases (GPX7 and GPX8) are key enzymes to reduce hydrogen peroxide in the ER. When this oxido-reductive system does not work properly, abnormal accumulation of misfolded proteins occurs in the ER and a set of signals named the unfolded protein response (UPR) is transmitted to the cytoplasm and nucleus to reestablish the ER homeostasis [43]. Three membrane-associated proteins have been identified for sensing ER stress in eukaryotes: activating transcription factor 6 (ATF6), pancreatic ER eIF2? kinase (PERK, also double-stranded RNA-activated protein kinase-like ER kinase), and inositol-requiring kinase1 (IRE1). The luminal domain of each sensor is bound to a 78 kDa chaperone termed glucose-regulated protein (GRP78/BIP). BIP dissociates upon ER stress to bind unfolded proteins, leading to the activation of the three sensors [44].
NRF2 and its homologue NRF1, also related to the antioxidant response, participate in the transduction of the UPR to the nucleus. In the case of NRF1, this protein is located at the ER membrane and undergoes nuclear translocation upon deglycosylation or cleavage. Then, UPR activation leads to the processing of NRF1 and nuclear accumulation of the resulting fragment in the nuclear compartment. However, the ability to transactivate ARE-containing genes of this NRF1 fragment is still under discussion [45].
Glover-Cutter and co-workers showed activation of the NRF2 orthologue of C. elegans, SKN-1, with different ER stressors. Increased SKN-1 expression was dependent on different UPR mediators, including IRE1 or PERK worm orthologues [46]. In PERK-deficient cells, impaired protein synthesis leads to accumulation of endogenous peroxides and subsequent apoptosis [47]. The effector used by PERK to protect the ER from these peroxides might be NRF2, since it has been reported that PERK phosphorylates NRF2 at Ser40, thus preventing its degradation by KEAP1 [31]. The induction of ASK1 is also likely to play a role in this route through the TRAF2-mediated kinase action of IRE1 [48]. Although the role of MAPKs in the regulation of NRF2 is still controversial, it was recently suggested that the IRE1-TRAF2-ASK1-JNK pathway might activate NRF2 [49] (Fig. 1). Interestingly, in C. elegans and human cells, new evidence suggests that cysteine sulfenylation of IRE1 kinase at its activation loop inhibits IRE1-mediated UPR and initiates a p38 antioxidant response driven by NRF2. The data suggest that IRE1 has an ancient function as a cytoplasmic sentinel that activates p38 and NRF2 [50].
Figure 1 Regulation of NRF2 by the UPR. Accumulation of unfolded or misfolded proteins inside the endoplasmic reticulum can initiate the unfolded protein response (UPR). First, the chaperone BIP is released from the intraluminal domain of the ER sensors IRE1 and PERK to bind unfolded/misfolded proteins. This enables dimerization and trans-auto-phosphorylation of their cytosolic domains. PERK activation results in direct NRF2 phosphorylation at Ser40, leading to NRF2 translocation to the nucleus and activation of target genes. IRE1 activation induces the recruitment of TRAF2 followed by ASK1 and JNK phosphorylation and activation. As JNK has been reported to phosphorylate and activate NRF2, it is reasonable to think that IRE1 activation would lead to increased NRF2 activity.
Many studies on the induction of the UPR have been conducted with the inhibitor of protein glycosylation tunicamycin. NRF2 appears to be essential for prevention of tunicamycin-induced apoptotic cell death [31] and its activation under these conditions is driven by the autophagic degradation of KEAP1 [51]. Accordingly, shRNA-mediated silencing of NRF2 expression in ?TC-6 cells, a murine insulinoma ?-cell line, significantly increased tunicamycin-induced cytotoxicity and led to an increase in the expression of the pro-apoptotic ER stress marker CHOP10. On the other hand, NRF2 activation by 1,2-dithiole-3-thione (D3T) reduced tunicamycin cytotoxicity and attenuated the expression of CHOP10 and PERK [52]. Interestingly, olfactory neurons submitted to systemic application of tunicamycin increased NRF2 in parallel with other UPR-members such as CHOP, BIP, XBP1 [53]. These results have been extended to in vivo studies, as lateral ventricular infusion of tunicamycin in rats induced expression of PERK and NRF2 in the hippocampus accompanied by significant cognitive deficits, increased TAU phosphorylation and A?42 deposits [54].
NRF2 Up-Regulates Key Genes for the Maintenance of the ER Physiology
The ER lumen needs an abundant supply of GSH from the cytosol in order to maintain disulfide chemistry. NRF2 modulates crucial enzymes of the GSH metabolism in the brain, such as cystine/glutamate transport, ?-glutamate cysteine synthetase (?-GS), glutamate-cysteine ligase catalytic and modulator subunits (GCLC and GCLM), glutathione reductase (GR) and glutathione peroxidase (GPX) (reviewed in [55]). The relevance of NRF2 in the maintenance of GSH in the ER is supported by the finding that pharmacological or genetic activation of NRF2 results in increased GSH synthesis via GCLC/GCLM, while inhibiting the expression of these enzymes by NRF2-knockdown caused an accumulation of damaged proteins within the ER leading to the UPR activation [56].
In C. elegans several components of the UPR target genes regulated by SKN-1, including Ire1, Xbp1 and Atf6. Although NRF2 up-regulates the expression of several peroxidase (PRX) and glutathione peroxidase (GPX) genes in mammals (reviewed in [57]), only GPX8 is a bona fide ER-localized enzyme, harboring the KDEL retrieval signal [58]. Loss of GPX8 causes UPR activation, leakage of ERO1?-derived hydrogen peroxide to the cytosol and cell death. Hydrogen peroxide derived from ERO1? activity cannot diffuse from the ER to the cytosol owing to the concerted action of GPX8 and PRX4 [59]. In this regard, an analysis of the antioxidant defense pathway-gene expression array using RNA from wild type and NRF2-null mice tissue, revealed that the expression of GPX8 was down-regulated in the absence of NRF2 [60]. In line with this, transcriptome analysis from patient samples suffering from myeloproliferative neoplasms, polycythemia or myelofibrosis, diseases also associate with oxidative stress and low-grade chronic inflammation, show lower expression levels of both NRF2 and GPX8 compared with control subjects [61]. There are not yet studies that specifically involve GPX8 in human brain protection but a transcriptome analysis in mice indicates a compensatory GPX8 increase in response to the Parkinsonian toxin MPTP [62].
Impact of NRF2 on the UPR Dysregulation in Neurodegenerative Diseases
Malfunction of PDI enzymes and chronic activation of the UPR might subsequently initiate or accelerate neurodegeneration. Disease-affected neurons, animal models of neurodegenerative disease as well as post-mortem human tissues evidenced up-regulation of several UPR-markers in most of these disorders. The alteration of PDI/UPR pathway in neurodegenerative diseases has been nicely reviewed in [63] but the following highlights from brain post-mortem samples should be considered. PDI levels are increased in tangle-bearing neurons and in Lewy Bodies of AD and PD patients, respectively [64], [65]. PDI and ERP57 are up-regulated in CSF from ALS patients and in brains from CJD subjects [66], [67], [68]. BIP, PERK, IRE1 and ATF6 are elevated in samples from patients with AD, PD or ALS [69], [70], [71], [67]. BIP, CHOP and XBP1 are elevated in post-mortem brain samples from HD [72], [73]. Moreover, up-regulation of ERP57, GRP94 and BIP was found in cortex tissues from CJD patients [74]. Altogether, this evidence reveals that the accumulation of misfolded proteins in the brain parenchyma leads to a deleterious and chronic activation of the UPR. Interestingly, there is a recent study linking activation of NRF2 by PERK in early AD. In this study, the authors analyzed whether oxidative stress mediated changes in NRF2 and the UPR may constitute early events in AD pathogenesis by using human peripheral blood cells and an AD transgenic mouse model at different disease stages. Increased oxidative stress and increased pSer40-NRF2 were observed in human peripheral blood mononuclear cells isolated from individuals with mild cognitive impairment. Moreover, they reported impaired ER calcium homeostasis and up-regulated ER-stress markers in these cells from individuals with mild cognitive impairment and mild AD [75].
Mutual Regulation of NRF2 and the Ubiquitin Proteasome�System (UPS)
The UPS Modulates NRF2 Protein Levels
The UPS participates in the degradation of damaged or misfolded proteins and controls the levels of key regulatory molecules in the cytosol and the nucleus. The central core of this system is a large multisubunit enzyme that contains a proteolytic active complex named 20S. The 20S core proteasome degrades unfolded proteins, but binding to different regulatory protein complexes changes its substrate specificity and activity. For instance, the addition of one or two 19S regulatory subunits to the 20S core constitutes the 26S proteasome and changes its specificity towards native folded proteins [76], [77]. Proteasomal degradation needs covalent binding of ubiquitin. Conjugation of ubiquitin proceeds via a three-step cascade mechanism. First, the ubiquitin-activating enzyme E1 activates ubiquitin in an ATP-requiring reaction. Then, one E2 enzyme (ubiquitin-carrier protein or ubiquitin-conjugating enzyme) transfers the activated ubiquitin from E1 to the substrate that is specifically bound to a member of the ubiquitin-protein ligase family, named E3. Although the exact fate of the ubiquitinated-protein will depend on the nature of the ubiquitin chain, this process generally results in the degradation by the 26S proteasome [78].
The E3-ligase KEAP1 is the best known inhibitor of NRF2. The mechanism of KEAP1 regulation elegantly explains how NRF2 levels adjust to oxidant fluctuations. Under basal conditions, newly synthesized NRF2 is grabbed by the homodimer KEAP1, which binds one NRF2 molecule at two amino acid sequences with low (aspartate, leucine, glycine; DLG) and high (glutamate, threonine, glycine, glutamate; ETGE) affinity. The interaction with KEAP1 aids to present NRF2 to the CULLIN3/RBX1 protein complex, resulting in its ubiquitination and subsequent proteasomal degradation. However, redox modification of KEAP1 impedes presentation of NRF2 to the UPS represented by CULLIN3/RBX1. As a result, newly synthetized NRF2 escapes KEAP1-dependent degradation, accumulates in the nucleus and activates ARE-containing genes [79], [80], [81], [82].
The E3-ligase adaptor ?-TrCP is also a homodimer that participates in the signaling events related to the phosphorylation of NRF2 by GSK-3?. This kinase phosphorylates specific serine residues of NRF2 (aspartate, serine, glycine, isoleucine serine; DSGIS) to create a degradation domain that is then recognized by ?-TrCP and tagged for proteasome degradation by a CULLIN1/RBX1 complex. The identification of the specific amino acids that are phosphorylated by GSK-3? in this degron was conducted by a combination of site-directed mutagenesis of the Neh6 domain, 2D-gel electrophoresis [15], [26] and mass spectroscopy [83]. Consequently, inhibition of GSK-3? by highly selective drugs or siRNAs against GSK-3 isoforms resulted in an increase in NRF2 protein levels. Similar results were found with siRNAs against ?-TrCP isoforms 1 and 2. Stabilization of NRF2 following GSK-3? inhibition occurred in KEAP1-deficient mouse embryo fibroblasts and in an ectopically expressed NRF2 deletion mutant lacking the critical ETGE residues for high-affinity binding to KEAP1, further demonstrating a KEAP1-independent regulation.
In the context of neurodegenerative diseases, we can envision the modulation of NRF2 by the UPS in two different ways. On the one hand, the KEAP1 system would sense redox imbalance derived from misfolded protein accumulation, while GSK-3/?-TrCP axis would act as an active participant in signaling transduction altered by loss of proteostasis (Fig. 2).
Figure 2 The UPS tightly controls NRF2 levels. Under homeostatic conditions, low NRF2 levels are maintained by the action of the E3 ligases adaptors KEAP1 and ?-TrCP. Left, NRF2 binds to the Kelch domains of a KEAP1 homodimer through a low (DLG) and a high (ETGE) affinity motifs. Through its BTB domain, KEAP1 simultaneously binds to a CULLIN3/RBX1 complex, enabling NRF2 ubiquitination and degradation by the 26 S proteasome. Moreover, GSK-3? phosphorylates Ser335 and Ser338 residues of NRF2 to create a degradation domain (DpSGIpSL) that is then recognized by the ubiquitin ligase adaptor ?-TrCP and tagged for proteasome degradation by a CULLIN3/RBX1 complex. Right, Upon exposure to reactive oxygen species or electrophiles critical Cys residues in KEAP1 are modified, rendering KEAP1 unable of interacting efficiently with NRF2 or CULLIN3/RBX1 and then this transcription factor increases its half-life and transcriptional activity towards ARE-genes. Signaling pathways that result in inhibition of GSK-3?, such AKT phosphorylation at Ser9, result in NRF2 impaired degradation by the proteasome, accumulation and induction of target genes.
NRF2 Increases UPS Activity Through the Transcriptional Control of Proteasome Subunits
NRF2 up-regulates the expression of several proteasome subunits, thus protecting the cell from the accumulation of toxic proteins. Twenty proteasome- and ubiquitination-related genes appear to be regulated by NRF2, according to a wide microarray analysis from liver RNA that was set up with the NRF2 inducer D3T [84]. In a posterior study, the same authors evidenced that the expression of most subunits of the 26S proteasome were enhanced up to three-fold in livers from mice treated with D3T. Subunit protein levels and proteasome activity were coordinately increased. However, no induction was seen in mice where the transcription factor NRF2 was disrupted. Promoter activity of the PSMB5 (20S) proteasome subunit increased with either NRF2 overexpression or treatment with activators in mouse embryonic fibroblasts, and AREs were identified in the proximal promoter of PSMB5 [85]. Pharmacological activation of NRF2 resulted in elevated expression levels of representative proteasome subunits (PSMA3, PSMA6, PSMB1 and PSMB5) only in non-senescent human fibroblasts containing functional NRF2 [86]. NRF2 activation during adaptation to oxidative stress results in high expression of the PSMB1 (20S) and PA28? subunits (or S11, proteasome regulator) [87]. Moreover, results from human embryonic stem cells revealed that NRF2 controls the expression of the proteasome maturation protein (POMP), a proteasome chaperone, which in turn modulates the proliferation of self-renewing human embryonic stem cells, three germ layer differentiation and cellular reprogramming [88]. All together, these studies indicate that NRF2 up-regulates the expression of key components of the UPS and therefore actively contributes to the clearance of proteins that otherwise would be toxic.
The NRF2-UPS Axis in Neurodegenerative Diseases
The role of the UPS in neurodegenerative diseases is a field of intensive debate. Initial studies reported decreased proteasome activity in human necropsies of patients affected from several neurodegenerative diseases. However, other studies employing in vitro and in vivo approaches found unchanged or even increased proteasome activity (reviewed in [89]). One possible explanation for this discrepancy is that the levels of the UPS components might change during disease progression and in different brain regions as is has been suggested for NRF2-targets.
Despite this controversy, it should be noted that up-regulation of ARE-containing proteasome genes will reinforce the UPS by increasing the clearance of toxic proteins in the brain. Indeed, ablation of NRF1, also modulator of the antioxidant response, in neuronal cells leads to impaired proteasome activity and neurodegeneration. Chromatin immunoprecipitation experiments and transcriptional analysis demonstrated that PSMB6 is regulated by NRF1. In addition, gene expression profiling led to the identification of NRF1 as a key transcriptional regulator of proteasome genes in neurons, suggesting that perturbations in NRF1 may contribute to the pathogenesis of neurodegenerative diseases [90]. Interestingly, NRF1 and its long isoform called TCF11 were shown to up-regulate ARE-containing proteasome genes upon proteasome inhibition in a feedback loop to compensate for reduced proteolytic activity [91], [92].
Regarding NRF2, there is a correlation among reduction of NRF2, RPT6 (19 S) and PSMB5 (20 S) levels in the midbrain of DJ-1-deficient mice treated with the neurotoxin paraquat [93]. Moreover, the naturally-occurring compound sulforaphane (SFN) gives a more robust image of NRF2 as a crucial modulator of the UPS. In vitro experiments with murine neuroblastoma Neuro2A cells evidenced an enhanced expression of the catalytic subunits of the proteasome, as well as its peptidase activities in response to SFN. This drug protected cells from hydrogen peroxide-mediated cytotoxicity and protein oxidation in a manner dependent on proteasome function [94]. In addition, Liu and co-workers employed a reporter mouse to monitor the UPS activity in response to SFN in the brain. These mice ubiquitously express the green fluorescence protein (GFP) fused to a constitutive degradation signal that promotes its rapid degradation by the UPS (GFPu). In cerebral cortex, SFN reduced the level of GFPu with a parallel increase in chymotrypsin-like (PSMB5), caspase-like (PSMB2), and trypsin-like (PSMB1) activities of the 20 S proteasome. In addition, treatment of Huntington-derived cells with SFN revealed that NRF2 activation enhanced mHtt degradation and reduced mHtt cytotoxicity [95]. The major mechanism of SFN action is through induction of NRF2 [96]. The specific contribution of NRF2 should be addressed employing NRF2-null systems in further studies.
Functional Connection Between NRF2 and Macroautophagy
NRF2 Protein Levels are Modulated by the Adaptor Protein P62
Autophagy refers to the degradation of cytosolic components inside lysosomes. This process is used for the clearance of long-lived and misfolded proteins as well as damaged organelles. A direct link between NRF2 and autophagy was first observed in connection with the adaptor protein p62, also termed SQSTM1 [97], [98], [99], [100], [101]. This protein shuttles ubiquitinated proteins to the proteasomal and lysosomal degradation machineries and sequesters damaged proteins into aggregates prior to their degradation. P62 presents an ubiquitin-associated (UBA) domain, for binding to ubiquitinated proteins, and a LC3-interacting region (LIR) for integration with the autophagosomal membrane through the autophagy receptor LC3.
Although the p62-mediated induction of NRF2 and its target genes was first reported in 2007 [102], the molecular mechanism was not fully understood until the discovery of its interaction with KEAP1 [103], [98], [99], [100], [101]. Komatsu and coworkers identified a KEAP1 interacting region (KIR) in p62 that bound KEAP1 in the same basic surface pocket as NRF2 and with a binding affinity similar to the ETGE motif in NRF2, suggesting competition between p62 and NRF2. The phosphorylation of Ser351 in the KIR motif in p62 (349-DPSTGE-354) was shown to increase its affinity for KEAP1, competing with NRF2 binding and allowing its accumulation and transcriptional activation of its target genes [98], [99]. In fact, p62 overexpression led to reduced NRF2 ubiquitination and consequent stabilization as well as induction of its target genes [104]. Some kinases have been suggested to participate in p62 phosphorylation. The mammalian target of rapamycin complex 1 (mTORC1) may be implicated, as treatment with the mTOR inhibitor rapamycin suppressed the phosphorylation of p62 and the down-regulation of KEAP1 upon arsenite treatment. Recently, it was demonstrated that TGF-?-activated kinase 1 (TAK1) could also phosphorylate p62, enhancing KEAP1 degradation and NRF2-up-regulation. The authors of this study suggest this is a way to regulate cellular redoxtasis under steady-state conditions, as TAK1-deficiency up-regulates ROS in the absence of any exogenous oxidant in different mouse tissues in parallel with a reduction in NRF2 protein levels [105].
A p62 construct lacking the UBA domain was still capable of binding KEAP1, implying that the interaction did not depend on ubiquitinated KEAP1 [101]. However, the p62 homologue in Drosophila melanogaster, named Ref(2), does not contain a KIR motif and does not directly interact with DmKEAP1, although it can bind to ubiquitinated DmKEAP1 through the UBA domain. Moreover, DmKEAP1 can directly interact with Atg8 (homologue to mammalian LC3). KEAP1 deficiency results in Atg8 and autophagy induction dependent on the NRF2 orthologue CncC and independent on TFEB/MITF [106]. The relationship between NRF2 and autophagy seems to be conserved though, highlighting its functional relevance.
The induction of NRF2 by p62 is the result of both the competition to bind KEAP1 and degradation of KEAP1 in the lysosome. Silencing of p62 with siRNA doubled KEAP1 half-life in parallel with a decrease in NRF2 and its target genes [101]. In agreement, ablation of p62 expression evidenced increased levels of KEAP1 compared with wild type mice. Very relevant, the increment in KEAP1 levels was not affected by proteasome inhibitors but was reduced under starvation-inducing autophagy [107]. In fact, KEAP1 is present in mammalian cells in autophagic vesicles decorated with p62 and LC3 [99], [100], [103]. All these data suggest that KEAP1 is a substrate of the macroautophagy machinery, but this issue should be analyzed with more detail because of the existence of some controversial results. KEAP1 protein levels were increased in Atg7-null mice, a key effector of macroautophagy [107], but pharmacological inhibition of macroautophagy with torin1, E64/pepstatin or bafilomycin failed to accumulate KEAP1 [107], [100]. Overall, these results suggest that increased p62 levels sequester KEAP1 into autophagic vacuoles and probably these results in KEAP1 autophagic degradation allowing NRF2 activation (Fig. 3). Two different studies reported that the sulfinic acid reductases SESTRINS play an important role in this context. SESTRIN 2 interacts with p62, KEAP1 and RBX1 and facilitates p62-dependent degradation of KEAP1 and NRF2 activation of target genes [108]. Another study showed that SESTRIN 2 interacted with ULK1 and p62, promoting phosphorylation of p62 at Ser403 which facilitated degradation of cargo proteins including KEAP1 [109].
Figure 3 NRF2 levels are regulated by the adaptor protein p62. The phosphorylation of Ser 351 in the KIR motif of p62 (349-DPSTGE-354) by mTORC1, TAK1 or other kinases results in increased affinity for binding to KEAP1 due to resemblance to the ETGE motif in NRF2. As a consequence, phosphorylated p62 displaces NRF2 and binds KEAP1. The LIR motif in p62 enables interaction with LC3 in the autophagosomal membrane, so that p62-KEAP1 complex is eventually degraded in the lysosome. As a consequence NRF2 is able to accumulate, translocate to the nucleus and increase the transcription of ARE-containing genes, including p62. This regulatory mechanism provides a perdurable NRF2 response, as KEAP1 has to be newly synthesized in order to inhibit NRF2 activity.
Modulation of Macroautophagy Genes by NRF2
NRF2 regulates the expression of relevant genes for macroautophagy as well as it does for the UPR and the UPS. The first evidence came from studies in which p62 expression was shown to be induced upon exposure to electrophiles, ROS and nitric oxide [110], [111], [112]. The mechanism of induction was described some years later with the finding that p62 contains a functional ARE in its gene promoter [99]. In a recent study, several other functional AREs were found and validated following bioinformatics analysis and ChIP assays. Moreover, mouse embryonic fibroblasts and cortical neurons from Nrf2-knockout mice exhibited reduced p62 expression, which could be rescued with an NRF2-expressing lentivirus. Similarly, NRF2 deficiency reduced p62 levels in injured neurons from mice hippocampus [36]. Therefore, it has been suggested that NRF2 activation increases p62 levels, resulting in KEAP1 degradation and favoring further NRF2 stabilization in a positive feedback loop. This non-canonical mechanism of NRF2 induction requires changes in gene expression and might be a relevant response to prolonged cellular stress.
The cargo recognition protein NDP52 was shown to be transcriptionally regulated by NRF2. NDP52 works in a similar way to p62, recognizing ubiquitinated proteins and interacting with LC3 through a LIR domain, so that cargoes are degraded in lysosomes. Five putative AREs were found in Ndp52 promoter DNA sequence. Three of them were identified with different mutant constructs and ChIP assays as indispensable for NRF2-mediated Ndp52 transcription [113]. Of note, Ndp52 mRNA levels were reduced in the hippocampus of Nrf2-knockout mice. One of these sequences was also validated in an independent study as an NRF2-regulated ARE [36].
However, the role of NRF2 in the modulation of autophagy is not limited to the induction of these two cargo-recognition proteins. In order to gain deeper insight in the role of NRF2 in the modulation of additional autophagy-related genes, our group screened the chromatin immunoprecipitation database ENCODE for two proteins, MAFK and BACH1, which bind the NRF2-regulated AREs. Using a script generated from the JASPAR’s consensus ARE sequence, we identified several putative AREs in many autophagy genes. Twelve of these sequences were validated as NRF2 regulated AREs in nine autophagy genes, whose expression was diminished in mouse embryo fibroblasts of Nrf2-knockout mice but could be restored by an NRF2-expressing lentivirus. Our study demonstrated that NRF2 activates the expression of some genes involved in different steps of the autophagic process, including autophagy initiation (ULK1), cargo recognition (p62 and NDP52), autophagosome formation (ATG4D, ATG7 and GABARAPL1), elongation (ATG2B and ATG5), and autolysosome clearance (ATG4D). Consequently, autophagy flux in response to hydrogen peroxide was impaired when NRF2 was absent [36].
Relevance of NRF2-Mediated Macroautophagy Genes Expression in Neurodegenerative Disorders
Defective autophagy has been shown to play an important role in several neurodegenerative diseases [114] and ablation of autophagy leads to neurodegeneration in mice [115], [116]. Atg7-knockout mice revealed that autophagy deficiency results in p62 accumulation in ubiquitin-positive inclusion bodies. KEAP1 was sequestered in these inclusion bodies, leading to NRF2 stabilization and induction of target genes [103]. Importantly, excessive accumulation of p62 together with ubiquitinated proteins has been identified in neurodegenerative diseases, including AD, PD and ALS [117]. In fact, neurons expressing high levels of APP or TAU of AD patients also expressed p62 and nuclear NRF2, suggesting their attempt to degrade intraneuronal aggregates through autophagy [36].
NRF2 deficiency aggravates protein aggregation in the context of AD. In fact, increased levels of phosphorylated and sarkosyl-insoluble TAU are found in Nrf2-knockout mice, although no difference in kinase or phosphatase activities could be detected comparing with the wild-type background [113]. Importantly, NDP52 was demonstrated to co-localize with TAU in murine neurons and direct interaction between phospho-TAU and NDP52 was shown by co-immunoprecipitation experiments both in mice and AD samples, pointing to its role in TAU degradation. Interestingly, silencing of NDP52, p62 or NRF2 in neurons resulted in increased phospho-TAU [113], [118]. Moreover, increased intraneuronal APP aggregates were found in the hippocampus of APP/PS1?E9 mice when NRF2 was absent. This correlated with altered autophagy markers, including increased phospho-mTOR/mTOR and phospho-p70S6k/p70S6k ratios (indicative of autophagy inhibition), augmented levels of pre-cathepsin D and a larger number of multivesicular bodies [119]. In mice co-expressing human APP (V717I) and TAU (P301L), NRF2 deficiency led to increased levels of total and phospho-TAU in the insoluble fraction and increased intraneuronal APP aggregates, together with reduced neuronal levels of p62, NDP52, ULK1, ATG5 and GABARAPL1. Co-localization between the adaptor protein p62 and APP or TAU was reduced in the absence of NRF2 [36]. Overall, these results highlight the importance of NRF2 in neuronal autophagy.
Different Transcription Factors Act Coordinately to Modulate Proteostasis
Under steady state conditions, proteostasis is controlled via protein-protein interactions and post-translational modifications obtaining a rapid response. However, cellular adaptation requires the transcriptional regulation of the UPR, UPS and autophagy genes. Considering that nerve cells are continuously submitted to low-grade toxic insults, including oxidative and proteotoxic stress, a reinforcement of proteostasis induced by transcriptional modulation might help preventing brain degeneration.
In the case of the UPR, the activation of each of the three arms will finally result in the transcriptional induction of certain genes (reviewed in [43]). For instance, an ATF6-derived fragment (ATF6f) binds to ER-stress response elements (ERSE) and induces the expression of several genes, including XBPI, BIP and CHOP. In addition, PERK signaling leads to the activation of the transcription factor ATF4, which controls the expression of multiple UPR-related genes and some others including the NRF2 target genes Hmox1 and p62. Finally, IRE1 activation results in the generation of an active transcription factor, spliced XBP1 (XBP1s), which controls the transcription of genes encoding proteins involved in protein folding.
On the other hand, NRF1 was shown to be necessary for proteasomal gene expression in the brain, as Nrf1-knockout mice exhibited reduced expression of genes encoding various subunits of the 20S core, as well the 19S regulatory complex together with impaired proteasomal function [90]. Both NRF1 and NRF2 bind to ARE sequences in the promoter regions of its target genes, which suggests they have overlapping transcriptional activities, although they differ in their regulatory mechanisms and cellular localization [120].
Transcription factors of the Forkhead box O (FOXO) family control the expression of multiple autophagy-related genes. Similar to what occurs with NRF2, there are multiple layers of regulation of the activity of FOXO members, which can be induced upon nutritional or oxidative stress [121]. Finally, the transcription factor TFEB, considered the master regulator of lysosomal biogenesis, plays a crucial role in regulation of autophagy under nutritional stress conditions. Thus, inhibition of mTORC1 leads to nuclear translocation of TFEB and induction of the expression of autophagy genes [122].
Overall, the existence of different transcriptional regulators of these machineries also suggests crosstalk and partially redundant mechanisms that may assure proteostasis under different circumstances. Accordingly, NRF2 may have a relevant role in tissues that support high levels of oxidative stress. For instance, oxidative stress-induced NRF2 may function under nutrient-rich conditions to transcriptionally up-regulate autophagy, similar to what has been found for TFEB under starvation conditions. Moreover, the brain functions largely under nutrient-rich conditions, posing NRF2 as a relevant mechanism to activate autophagy in neurons.
Promising�Therapeutic Potential for NRF2 in Proteinopathies
In the past few years, a great progress has been made in the knowledge of the regulatory roles of the UPR, UPS and autophagy on NRF2 activity, as well as the reciprocal NRF2-mediated transcription of components of these three systems. Therefore, new therapeutic possibilities may arise based on the exploitation of NRF2 as a crucial regulator of protein clearance in neurodegenerative diseases.
However, a key remaining question is whether it will be useful or deleterious to increase NRF2 levels in brain. Analysis of epidemiological data may provide a partial answer, as it indicates that the NFE2L2 gene is highly polymorphic and some single nucleotide polymorphisms found in its promoter regulatory region may provide a range of �physiological� variability in gene expression at the population level and some haplotypes were associated with decreased risk and/or delayed onset of AD, PD or ALS [123]. Moreover, as discussed by Hayes and colleagues [124], NRF2 effect might have an U-shaped response, meaning that too low NRF2 levels may result in a loss of cytoprotection and increased susceptibility to stressors, while too much NRF2 might disturb homeostatic balance towards a reductive scenario (reductive stress), which would favor protein misfolding and aggregation. Low NRF2 levels in the brain support the idea that a slight up-regulation may be sufficient to achieve a benefit under pathological conditions. In fact, the protective role of pharmacological NRF2-mediated activation of protein clearance has been shown in different neurodegeneration cell culture and in vivo models.
SFN is a pharmacological NRF2 activator that was demonstrated to induce proteasomal and autophagy gene expression [95], [36]. Interestingly, Jo and colleagues demonstrated that SFN reduced the levels of phosphorylated TAU and increased Beclin-1 and LC3-II, suggesting NRF2 activation may facilitate degradation of this toxic protein through autophagy [113]. Moreover, degradation of mHtt was enhanced with SFN, and this was reverted with the use of MG132, indicating proteasomal degradation of this toxic protein [95]. Autophagy-mediated degradation of phospho- and insoluble-TAU was reported with the organic flavonoid fisetin. This compound was able to induce autophagy by simultaneously promoting the activation and nuclear translocation of both TFEB and NRF2, along with some of its target genes. This response was prevented by TFEB or NRF2 silencing [125]. Bott and colleagues reported beneficial effects of a simultaneous NRF2, NRF1 and HSF1 activator on protein toxicity in spinal and bulbar muscular atrophy, a neurodegenerative disorder caused by expansion of polyglutamine-encoding CAG repeats in which protein aggregates are present [126]. The potential of NRF2 activation for the treatment of neurodegenerative disorders has been demonstrated with the approval of BG-12, the oral formulation of the NRF2 inducer dimethyl fumarate (DMF), for the treatment of multiple sclerosis [127], [128]. The success of DMF with autoimmune diseases with a strong inflammatory component suggests that neurodegenerative diseases might benefit from repositioning this drug. In a recent preclinical study of an ?-synucleinopathy model of PD, DMF was shown to be neuroprotective due, in part, to its induction of autophagy [129]. Studies reporting beneficial effects of NRF2 on neurodegeneration but not focusing on its effect on protein clearance are even more abundant (for a comprehensive review, see [7]). This is quite relevant, as it highlights the multiple damaging processes that can be simultaneously targeted by a single hit in NRF2, also including oxidative stress, neuroinflammation or mitochondrial dysfunction. However, future work will be needed to definitely determine if pharmacological activation of NRF2 may be a valid strategy to facilitate degradation of toxic proteins in the brain.
As explained before, exacerbated GSK-3? activity was reported in neurodegenerative diseases and it has been speculated that consequent NRF2 reduction can be partially responsible for the deleterious outcome. Under these pathological conditions, GSK-3 inhibitors could also cooperate to increase NRF2 levels and proteostasis. The beneficial effects of GSK-3 inhibitors have been reported in different models of neurodegeneration and, more interesting, GSK-3 repression was shown to reduce the levels of toxic proteins [130], [131], [132], [133]. Although no direct links between GSK-3 inhibition and NRF2-transcriptional regulation of genes promoting proteostasis have been observed yet, it is reasonable to speculate that down-regulation of GSK-3 activity would result in increased NRF2 levels, which eventually will result in reinforced proteostasis.
The transcriptional activity of NRF2 as well as the cellular capacity to maintain proteostasis decrease with age, the main risk factor for the development of neurodegenerative diseases. It is reasonable to think that the reinforcement of NRF2 and, consequently, proteostasis would, at least, delay the accumulation of protein aggregates and neurodegeneration. Indeed, treatment of human senescent fibroblasts with 18?-glycyrrhetinic acid (18?-GA) triterpenoid promoted NRF2 activation, leading to proteasome induction and enhanced life span. This study suggests that pharmacological activation of NRF2 is possible even in late life [86]. Moreover, a later study showed that this compound mediated SKN-1 and proteasome activation in C.elegans with beneficial effects on AD progression in relevant nematode models [134].
All things considered, NRF2-mediated induction of proteostasis-related genes seems to be beneficial in different proteinopathies.
Sulforaphane and Its Effects on Cancer, Mortality, Aging, Brain and Behavior, Heart Disease & More
Isothiocyanates are some of the most important plant compounds you can get in your diet. In this video I make the most comprehensive case for them that has ever been made. Short attention span? Skip to your favorite topic by clicking one of the time points below. Full timeline below.
Key sections:
00:01:14 – Cancer and mortality
00:19:04 – Aging
00:26:30 – Brain and behavior
00:38:06 – Final recap
00:40:27 – Dose
Full timeline:
00:00:34 – Introduction of sulforaphane, a major focus of the video.
00:01:14 – Cruciferous vegetable consumption and reductions in all-cause mortality.
00:02:12 – Prostate cancer risk.
00:02:23 – Bladder cancer risk.
00:02:34 – Lung cancer in smokers risk.
00:02:48 – Breast cancer risk.
00:03:13 – Hypothetical: what if you already have cancer? (interventional)
00:03:35 – Plausible mechanism driving the cancer and mortality associative data.
00:04:38 – Sulforaphane and cancer.
00:05:32 – Animal evidence showing strong effect of broccoli sprout extract on bladder tumor development in rats.
00:06:06 – Effect of direct supplementation of sulforaphane in prostate cancer patients.
00:07:09 – Bioaccumulation of isothiocyanate metabolites in actual breast tissue.
00:08:32 – Inhibition of breast cancer stem cells.
00:08:53 – History lesson: brassicas were established as having health properties even in ancient Rome.
00:09:16 – Sulforaphane’s ability to enhance carcinogen excretion (benzene, acrolein).
00:09:51 – NRF2 as a genetic switch via antioxidant response elements.
00:10:10 – How NRF2 activation enhances carcinogen excretion via glutathione-S-conjugates.
00:10:34 – Brussels sprouts increase glutathione-S-transferase and reduce DNA damage.
00:11:20 – Broccoli sprout drink increases benzene excretion by 61%.
00:13:31 – Broccoli sprout homogenate increases antioxidant enzymes in the upper airway.
00:15:45 – Cruciferous vegetable consumption and heart disease mortality.
00:16:55 – Broccoli sprout powder improves blood lipids and overall heart disease risk in type 2 diabetics.
00:19:04 – Beginning of aging section.
00:19:21 – Sulforaphane-enriched diet enhances lifespan of beetles from 15 to 30% (in certain conditions).
00:20:34 – Importance of low inflammation for longevity.
00:22:05 – Cruciferous vegetables and broccoli sprout powder seem to reduce a wide variety of inflammatory markers in humans.
00:36:32 – Sulforaphane improves learning in model of type II diabetes in mice.
00:37:19 – Sulforaphane and duchenne muscular dystrophy.
00:37:44 – Myostatin inhibition in muscle satellite cells (in vitro).
00:38:06 – Late-video recap: mortality and cancer, DNA damage, oxidative stress and inflammation, benzene excretion, cardiovascular disease, type II diabetes, effects on the brain (depression, autism, schizophrenia, neurodegeneration), NRF2 pathway.
00:40:27 – Thoughts on figuring out a dose of broccoli sprouts or sulforaphane.
00:41:01 – Anecdotes on sprouting at home.
00:43:14 – On cooking temperatures and sulforaphane activity.
00:43:45 – Gut bacteria conversion of sulforaphane from glucoraphanin.
00:44:24 – Supplements work better when combined with active myrosinase from vegetables.
00:44:56 – Cooking techniques and cruciferous vegetables.
00:46:06 – Isothiocyanates as goitrogens.
The nuclear factor erythroid-derived 2 (NF-E2)-related factor 2, otherwise known as Nrf2, is a transcription factor which regulates the expression of a variety of antioxidant and detoxifying enzymes. Research studies have also demonstrated its role in controlling oxidative stress. Most neurodegenerative diseases, such as Alzheimer’s disease and Parkinson’s disease, are characterized by oxidative stress and chronic inflammation, the common targets of Nrf2 treatment approaches. Dr. Alex Jimenez D.C., C.C.S.T. Insight
Concluding Remarks
Transcription factor NRF2 orchestrates a proteostatic response by sensing to and modulating changes in the UPR, UPS and autophagy (Fig. 4). Consequently, the lack of NRF2 has been shown to aggravate proteinopathy, suggesting that NRF2 is necessary for optimal protein clearance. All together, we can speculate that NRF2 might be an interesting therapeutic target for proteinopathies.
Figure 4 NRF2 as a hub connecting proteotoxic-derived emergency signals to a protective transcriptional response. The accumulation of unfolded/misfolded proteins will lead to the activation of the unfolded protein response (UPR) in the ER. Activation of PERK or MAPK may result in the transcriptional induction of the ER-resident Gpx8 and several enzymes regulating GSH levels, critical to ensure correct protein folding. Protein aggregates inhibit proteasome activity (UPS), probably avoiding NRF2 degradation. NRF2 has been shown to specifically modulate the transcription of Psma3, Psma6, Psmb1, Psmb5 and Pomp genes. Several other subunits were up-regulated in an NRF2-dependent manner in response to D3T, probably enlarging the list of proteasome subunits regulated by NRF2. Autophagy is the main pathway for the degradation of protein aggregates. Autophagy also regulates NRF2, connecting this degradation pathway with NRF2 transcriptional induction of p62, Ndp52, Ulk1, Atg2b, Atg4c, Atg5, Atg7 and Gabarapl1.
According to the article above, while the symptoms of neurodegenerative diseases can be treated through a variety of treatment options, research studies have demonstrated that Nrf2 activation can be a helpful treatment approach. Because Nrf2 activators target broad mechanisms of disease, all neurodegenerative diseases can benefit from the use of the Nrf2 transcription factor. The findings of Nrf2 have revolutionized the treatment of neurodegenerative diseases. The scope of our information is limited to chiropractic and spinal health issues. To discuss the subject matter, please feel free to ask Dr. Jimenez or contact us at�915-850-0900�.
Additional Topic Discussion: Relieving Knee Pain without Surgery
Knee pain is a well-known symptom which can occur due to a variety of knee injuries and/or conditions, including�sports injuries. The knee is one of the most complex joints in the human body as it is made-up of the intersection of four bones, four ligaments, various tendons, two menisci, and cartilage. According to the American Academy of Family Physicians, the most common causes of knee pain include patellar subluxation, patellar tendinitis or jumper’s knee, and Osgood-Schlatter disease. Although knee pain is most likely to occur in people over 60 years old, knee pain can also occur in children and adolescents. Knee pain can be treated at home following the RICE methods, however, severe knee injuries may require immediate medical attention, including chiropractic care. �
Oxidative stress is described as cell damage caused by free radicals, or unstable molecules, which can ultimately affect healthy function. The human body creates free radicals to neutralize bacteria and viruses, however, external factors, such as oxygen, pollution, and radiation, can often also produce free radicals. Oxidative stress has been associated with numerous health issues.
Oxidative stress and other stressors turn on internal protective mechanisms which can help regulate the human body’s antioxidant response. Nrf2 is a protein which senses levels of oxidative stress and enables the cells to protect themselves from internal and external factors. Nrf2 has also been demonstrated to help regulate genes involved in the production of antioxidant enzymes and stress-response genes. The purpose of the article below is to explain the effects of Nrf2 in cancer.
Abstract
The Keap1-Nrf2 pathway is the major regulator of cytoprotective responses to oxidative and electrophilic stress. Although cell signaling pathways triggered by the transcription factor Nrf2 prevent cancer initiation and progression in normal and premalignant tissues, in fully malignant cells Nrf2 activity provides growth advantage by increasing cancer chemoresistance and enhancing tumor cell growth. In this graphical review, we provide an overview of the Keap1-Nrf2 pathway and its dysregulation in cancer cells. We also briefly summarize the consequences of constitutive Nrf2 activation in cancer cells and how this can be exploited in cancer gene therapy.
The Keap1-Nrf2 pathway is the major regulator of cytoprotective responses to endogenous and exogenous stresses caused by reactive oxygen species (ROS) and electrophiles [1]. The key signaling proteins within the pathway are the transcription factor Nrf2 (nuclear factor erythroid 2-related factor 2) that binds together with small Maf proteins to the antioxidant response element (ARE) in the regulatory regions of target genes, and Keap1 (Kelch ECH associating protein 1), a repressor protein that binds to Nrf2 and promotes its degradation by the ubiquitin proteasome pathway (Fig. 1). Keap1 is a very cysteine-rich protein, mouse Keap1 having a total of 25 and human 27 cysteine residues, most of which can be modified in vitro by different oxidants and electrophiles [2]. Three of these residues, C151, C273 and C288, have been shown to play a functional role by altering the conformation of Keap1 leading to nuclear translocation of Nrf2 and subsequent target gene expression [3] (Fig. 1). The exact mechanism whereby cysteine modifications in Keap1 lead to Nrf2 activation is not known, but the two prevailing but not mutually exclusive models are (1) the �hinge and latch� model, in which Keap1 modifications in thiol residues residing in the IVR of Keap1 disrupt the interaction with Nrf2 causing a misalignment of the lysine residues within Nrf2 that can no longer be polyubiquitinylated and (2) the model in which thiol modification causes dissociation of Cul3 from Keap1 [3]. In both models, the inducer-modified and Nrf2-bound Keap1 is inactivated and, consequently, newly synthesized Nrf2 proteins bypass Keap1 and translocate into the nucleus, bind to the ARE and drive the expression of Nrf2 target genes such as NAD(P)H quinone oxidoreductase 1 (NQO1), heme oxygenase 1 (HMOX1), glutamate-cysteine ligase (GCL) and glutathione S transferases (GSTs) (Fig. 2). In addition to modifications of Keap1 thiols resulting in Nrf2 target gene induction, proteins such as p21 and p62 can bind to Nrf2 or Keap1 thereby disrupting the interaction between Nrf2 and Keap1 [1], [3] (Fig. 3).
Fig. 1. Structures of Nrf2 and Keap1 and the cysteine code. (A) Nrf2 consists of 589 amino acids and has six evolutionarily highly conserved domains, Neh1-6. Neh1 contains a bZip motif, a basic region � leucine zipper (L-Zip) structure, where the basic region is responsible for DNA recognition and the L-Zip mediates dimerization with small Maf proteins. Neh6 functions as a degron to mediate degradation of Nrf2 in the nucleus. Neh4 and 5 are transactivation domains. Neh2 contains ETGE and DLG motifs, which are required for the interaction with Keap1, and a hydrophilic region of lysine residues (7 K), which are indispensable for the Keap1-dependent polyubiquitination and degradation of Nrf2. (B) Keap1 consists of 624 amino acid residues and has five domains. The two protein�protein interaction motifs, the BTB domain and the Kelch domain, are separated by the intervening region (IVR). The BTB domain together with the N-terminal portion of the IVR mediates homodimerization of Keap1 and binding with Cullin3 (Cul3). The Kelch domain and the C-terminal region mediate the interaction with Neh2. (C) Nrf2 interacts with two molecules of Keap1 through its Neh2 ETGE and DLG motifs. Both ETGE and DLG bind to similar sites on the bottom surface of the Keap1 Kelch motif. (D) Keap1 is rich in cysteine residues, with 27 cysteines in human protein. Some of these cysteines are located near basic residues and are therefore excellent targets of electrophiles and oxidants. The modification pattern of the cysteine residues by electrophiles is known as the cysteine code. The cysteine code hypothesis proposes that structurally different Nrf2 activating agents affect different Keap1 cysteines. The cysteine modifications lead to conformational changes in the Keap1 disrupting the interaction between the Nrf2 DLG and Keap1 Kelch domains, thus inhibiting the polyubiquitination of Nrf2. The functional importance of Cys151, Cys273 and Cys288 has been shown, as Cys273 and Cys288 are required for suppression of Nrf2 and Cys151 for activation of Nrf2 by inducers [1], [3].
Fig. 2. The Nrf2-Keap1 signaling pathway. (A and B) in basal conditions, two Keap1 molecules bind to Nrf2 and Nrf2 is polyubiquitylated by the Cul3-based E3 ligase complex. This polyubiquitilation results in rapid Nrf2 degradation by the proteasome. A small proportion of Nrf2 escapes the inhibitory complex and accumulates in the nucleus to mediate basal ARE-dependent gene expression, thereby maintaining the cellular homeostasis. (C) Under stress conditions, inducers modify the Keap1 cysteines leading to the inhibition of Nrf2 ubiquitylation via dissociation of the inhibitory complex. (D) According to the hinge and latch model, modification of specific Keap1 cysteine residues leads to conformational changes in Keap1 resulting in the detachment of the Nrf2 DLG motif from Keap1. Ubiquitination of Nrf2 is disrupted but the binding with the ETGE motif remains. (E) In the Keap1-Cul3 dissociation model, the binding of Keap1 and Cul3 is disrupted in response to electrophiles, leading to the escape of Nrf2 from the ubiquitination system. In both of the suggested models, the inducer-modified and Nrf2-bound Keap1 is inactivated and, consequently, newly synthesized Nrf2 proteins bypass Keap1 and translocate into the nucleus, bind to the Antioxidant Response Element (ARE) and drive the expression of Nrf2 target genes such as NQO1, HMOX1, GCL and GSTs [1], [3].
Fig. 3. Mechanisms for constitutive nuclear accumulation of Nrf2 in cancer. (A) Somatic mutations in Nrf2 or Keap1 disrupt the interaction of these two proteins. In Nrf2, mutations affect ETGE and DLG motifs, but in Keap1 mutations are more evenly distributed. Furthermore, oncogene activation, such as KrasG12D[5], or disruption of tumor suppressors, such as PTEN [11] can lead to transcriptional induction of Nrf2 and an increase in nuclear Nrf2. (B) Hypermethylation of the Keap1 promoter in lung and prostate cancer leads to reduction of Keap1 mRNA expression, which increases the nuclear accumulation of Nrf2 [6], [7]. (C) In familial papillary renal carcinoma, the loss of fumarate hydratase enzyme activity leads to the accumulation of fumarate and further to succination of Keap1 cysteine residues (2SC). This post-translational modification leads to the disruption of Keap1-Nrf2 interaction and nuclear accumulation of Nrf2 [8], [9]. (D) Accumulation of disruptor proteins such as p62 and p21 can disturb Nrf2-Keap1 binding and results in an increase in nuclear Nrf2. p62 binds to Keap1 overlapping the binding pocket for Nrf2 and p21 directly interacts with the DLG and ETGE motifs of Nrf2, thereby competing with Keap1 [10].
Mechanisms of Activation and Dysregulation of Nrf2 in Cancer
Although cytoprotection provided by Nrf2 activation is important for cancer chemoprevention in normal and premalignant tissues, in fully malignant cells Nrf2 activity provides growth advantage by increasing cancer chemoresistance and enhancing tumor cell growth [4]. Several mechanisms by which Nrf2 signaling pathway is constitutively activated in various cancers have been described: (1) somatic mutations in Keap1 or the Keap1 binding domain of Nrf2 disrupting their interaction; (2) epigenetic silencing of Keap1 expression leading to defective repression of Nrf2; (3) accumulation of disruptor proteins such as p62 leading to dissociation of the Keap1-Nrf2 complex; (4) transcriptional induction of Nrf2 by oncogenic K-Ras, B-Raf and c-Myc; and (5) post-translational modification of Keap1 cysteines by succinylation that occurs in familial papillary renal carcinoma due to the loss of fumarate hydratase enzyme activity [3], [4], [5], [6], [7], [8], [9], [10] (Fig. 3). Constitutively abundant Nrf2 protein causes increased expression of genes involved in drug metabolism thereby increasing the resistance to chemotherapeutic drugs and radiotherapy. In addition, high Nrf2 protein level is associated with poor prognosis in cancer [4]. Overactive Nrf2 also affects cell proliferation by directing glucose and glutamine towards anabolic pathways augmenting purine synthesis and influencing the pentose phosphate pathway to promote cell proliferation [11] (Fig. 4).
Fig. 4. The dual role of Nrf2 in tumorigenesis. Under physiological conditions, low levels of nuclear Nrf2 are sufficient for the maintenance of cellular homeostasis. Nrf2 inhibits tumor initiation and cancer metastasis by eliminating carcinogens, ROS and other DNA-damaging agents. During tumorigenesis, accumulating DNA damage leads to constitutive hyperactivity of Nrf2 which helps the autonomous malignant cells to endure high levels of endogenous ROS and to avoid apoptosis. Persistently elevated nuclear Nrf2 levels activate metabolic genes in addition to the cytoprotective genes contributing to metabolic reprogramming and enhanced cell proliferation. Cancers with high Nrf2 levels are associated with poor prognosis because of radio and chemoresistance and aggressive cancer cell proliferation. Thus, Nrf2 pathway activity is protective in the early stages of tumorigenesis, but detrimental in the later stages. Therefore, for the prevention of cancer, enhancing Nrf2 activity remains an important approach whereas for the treatment of cancer, Nrf2 inhibition is desirable [4], [11].
Given that high Nrf2 activity commonly occurs in cancer cells with adverse outcomes, there is a need for therapies to inhibit Nrf2. Unfortunately, due to structural similarity with some other bZip family members, the development of specific Nrf2 inhibitors is a challenging task and only a few studies of Nrf2 inhibition have been published to date. By screening natural products, Ren et al. [12] identified an antineoplastic compound brusatol as an Nrf2 inhibitor that enhances the chemotherapeutic efficacy of cisplatin. In addition, PI3K inhibitors [11], [13] and Nrf2 siRNA [14] have been used to inhibit Nrf2 in cancer cells. Recently, we have utilized an alternative approach, known as cancer suicide gene therapy, to target cancer cells with high Nrf2 levels. Nrf2-driven lentiviral vectors [15] containing thymidine kinase (TK) are transferred into cancer cells with high ARE activity and the cells are treated with a pro-drug, ganciclovir (GCV). GCV is metabolized to GCV-monophosphate, which is further phosphorylated by cellular kinases into a toxic triphosphate form [16] (Fig. 5). This leads to effective killing of not only TK containing tumor cells, but also the neighboring cells due to the bystander effect [17]. ARE-regulated TK/GCV gene therapy can be further enhanced via combining a cancer chemotherapeutic agent doxorubicin to the treatment [16], supporting the notion that this approach could be useful in conjuction with traditional therapies.
Fig. 5. Suicide gene therapy. Constitutive Nrf2 nuclear accumulation in cancer cells can be exploited by using Nrf2-driven viral vector for cancer suicide gene therapy [16]. In this approach, lentiviral vector (LV) expressing thymidine kinase (TK) under minimal SV40 promoter with four AREs is transduced to lung adenocarcinoma cells. High nuclear Nrf2 levels lead to robust expression of TK through Nrf2 binding. Cells are then treated with a pro-drug, ganciclovir (GCV), which is phosphorylated by TK. Triphosphorylated GCV disrupts DNA synthesis and leads to effective killing of not only TK containing tumor cells, but also the neighboring cells due to the bystander effect.
Nrf2 is a master regulator which triggers the production of powerful antioxidants in the human body which help eliminate oxidative stress. Various antioxidant enzymes, such as superoxide dismutase, or SOD, glutathione, and catalase, are also activated through the Nrf2 pathway. Furthermore, certain phytochemicals like turmeric, ashwagandha, bacopa, green tea, and milk thistle, activate Nrf2. Research studies have found that Nrf2 activation can naturally enhance cellular protection and restore balance to the human body.
Dr. Alex Jimenez D.C., C.C.S.T. Insight
Sulforaphane and Its Effects on Cancer, Mortality, Aging, Brain and Behavior, Heart Disease & More
Isothiocyanates are some of the most important plant compounds you can get in your diet. In this video I make the most comprehensive case for them that has ever been made. Short attention span? Skip to your favorite topic by clicking one of the time points below. Full timeline below.
Key sections:
00:01:14 – Cancer and mortality
00:19:04 – Aging
00:26:30 – Brain and behavior
00:38:06 – Final recap
00:40:27 – Dose
Full timeline:
00:00:34 – Introduction of sulforaphane, a major focus of the video.
00:01:14 – Cruciferous vegetable consumption and reductions in all-cause mortality.
00:02:12 – Prostate cancer risk.
00:02:23 – Bladder cancer risk.
00:02:34 – Lung cancer in smokers risk.
00:02:48 – Breast cancer risk.
00:03:13 – Hypothetical: what if you already have cancer? (interventional)
00:03:35 – Plausible mechanism driving the cancer and mortality associative data.
00:04:38 – Sulforaphane and cancer.
00:05:32 – Animal evidence showing strong effect of broccoli sprout extract on bladder tumor development in rats.
00:06:06 – Effect of direct supplementation of sulforaphane in prostate cancer patients.
00:07:09 – Bioaccumulation of isothiocyanate metabolites in actual breast tissue.
00:08:32 – Inhibition of breast cancer stem cells.
00:08:53 – History lesson: brassicas were established as having health properties even in ancient Rome.
00:09:16 – Sulforaphane’s ability to enhance carcinogen excretion (benzene, acrolein).
00:09:51 – NRF2 as a genetic switch via antioxidant response elements.
00:10:10 – How NRF2 activation enhances carcinogen excretion via glutathione-S-conjugates.
00:10:34 – Brussels sprouts increase glutathione-S-transferase and reduce DNA damage.
00:11:20 – Broccoli sprout drink increases benzene excretion by 61%.
00:13:31 – Broccoli sprout homogenate increases antioxidant enzymes in the upper airway.
00:15:45 – Cruciferous vegetable consumption and heart disease mortality.
00:16:55 – Broccoli sprout powder improves blood lipids and overall heart disease risk in type 2 diabetics.
00:19:04 – Beginning of aging section.
00:19:21 – Sulforaphane-enriched diet enhances lifespan of beetles from 15 to 30% (in certain conditions).
00:20:34 – Importance of low inflammation for longevity.
00:22:05 – Cruciferous vegetables and broccoli sprout powder seem to reduce a wide variety of inflammatory markers in humans.
00:36:32 – Sulforaphane improves learning in model of type II diabetes in mice.
00:37:19 – Sulforaphane and duchenne muscular dystrophy.
00:37:44 – Myostatin inhibition in muscle satellite cells (in vitro).
00:38:06 – Late-video recap: mortality and cancer, DNA damage, oxidative stress and inflammation, benzene excretion, cardiovascular disease, type II diabetes, effects on the brain (depression, autism, schizophrenia, neurodegeneration), NRF2 pathway.
00:40:27 – Thoughts on figuring out a dose of broccoli sprouts or sulforaphane.
00:41:01 – Anecdotes on sprouting at home.
00:43:14 – On cooking temperatures and sulforaphane activity.
00:43:45 – Gut bacteria conversion of sulforaphane from glucoraphanin.
00:44:24 – Supplements work better when combined with active myrosinase from vegetables.
00:44:56 – Cooking techniques and cruciferous vegetables.
00:46:06 – Isothiocyanates as goitrogens.
Acknowledgments
This work was supported by the Academy of Finland, the Sigrid Juselius Foundation and the Finnish Cancer Organisations.
In conclusion, nuclear factor (erythroid-derived 2)-like 2, also known as NFE2L2 or Nrf2, is a protein which increases the production of antioxidants which protect the human body against oxidative stress. As described above, the stimulation of the Nrf2 pathway are being studies for the treatment of diseases caused by oxidative stress, including cancer. The scope of our information is limited to chiropractic and spinal health issues. To discuss the subject matter, please feel free to ask Dr. Jimenez or contact us at�915-850-0900�.
Additional Topic Discussion: Relieving Knee Pain without Surgery
Knee pain is a well-known symptom which can occur due to a variety of knee injuries and/or conditions, including�sports injuries. The knee is one of the most complex joints in the human body as it is made-up of the intersection of four bones, four ligaments, various tendons, two menisci, and cartilage. According to the American Academy of Family Physicians, the most common causes of knee pain include patellar subluxation, patellar tendinitis or jumper’s knee, and Osgood-Schlatter disease. Although knee pain is most likely to occur in people over 60 years old, knee pain can also occur in children and adolescents. Knee pain can be treated at home following the RICE methods, however, severe knee injuries may require immediate medical attention, including chiropractic care.
DNA supports approximately 20,000 genes, each holding a program for the creation of a protein or enzyme required for a healthy lifestyle. Every one of these patterns needs to be constantly regulated by a sort of “promoter” which manages exactly how much of each substance and/or chemical is generated and under which conditions these will also develop.
By connecting to a particular kind of the switch-like promoter areas, known as the Antioxidant Response Element, or ARE, the Nrf2 factor�supports the speed of creation for hundreds of distinct genes which enable the cells to survive under stressful circumstances. These genes then generate a selection of antioxidant enzymes which develop a defense network by neutralizing oxidants and by cleaning up the toxic by-products left behind in their�production, in addition to helping restore the�damage they caused.
What is Oxidative Stress?
Several oxidants like the superoxide radical, or O2-., and hydrogen peroxide, or H2O2, have been created through the practice of burning off the substances and/or chemicals which sustain the human body. The human body�possesses antioxidant enzymes which�neutralize and detoxify reactive foods and drinks we consume. The Nrf2 modulates their production to keep equilibrium and underscores the demand for all these enzymes. This balance can be interrupted by a�couple of factors, including age.
As we age,�the human body creates less Nrf2 and this delicate equilibrium can gradually begin to�turn towards the oxidative side, a state referred to as oxidative stress. Disease may also cause the overproduction of oxidants. Infections, allergies, and autoimmune disorders can additionally trigger our immune cells to create reactive oxidants, such as O2-. , H2O2, OH and HOCl, where healthy cells become damaged and respond with inflammation. Diseases associated with aging, including heart attacks, stroke, cancer, and neurodegenerative conditions like Alzheimer’s disease, also increase the development of oxidants, generating stress and an inflammation response.
What are Nrf2 Activators?
The Nrf2 protein, also called a transcription factor due to the way it can support and control enzymes and genes, is the secret element of a sequence of biochemical reactions within the cell which reacts to modifications in cognitive equilibrium as well as oxidative balance. The sensing elements of this pathway modify and discharge Nrf2, triggering it so it might spread into the nucleus of the cell towards the DNA. The Nrf2 may alternatively turn on or switch off the genes and enzymes it supports to protect the cell.
Fortunately, a variety of substances which are Nrf2 activators develop through the consumption of certain plants and extracts utilized centuries ago in Chinese and Native American traditional remedies. These phytochemicals seem to be equally as powerful with fewer side-effects, as the Nrf2-activating pharmaceutical products which are being used today.
Nuclear factor erythroid 2-related factor, more commonly known as Nrf2, is a transcription factor which protects the cell by regulating genes, enzymes and antioxidant responses. Transcription factors are a type of protein which attach to DNA to promote the creation of specific substances and chemicals, including glutathione S-transferases, or GSTs. Nrf2 activation induces the production of active proteins which exhibit a powerful antioxidant capacity to help decrease oxidative stress.
Dr. Alex Jimenez D.C., C.C.S.T. Insight
The Science Behind Nrf2 Activation
Once the initial Nrf2-activating dietary supplement was created in 2004, minimal information was known concerning the function of the Nrf2 pathway. Approximately 200 newspapers in the literature on Nrf2, also known as nuclear factor-like 2 or NFE2L2, existed and researchers were only just starting to discover the antioxidant response of Nrf2 in mammals. As of 2017, however, over 9,300 academic research studies on this “master regulator,” have been printed.
In reality, Nrf2 regulates many antioxidant enzymes which don’t correlate to the genes, instead, they offer protection against a variety of stress-related circumstances which are encountered by cells, organs and ultimately organisms, under healthy and pathological conditions. Based on this new quantity of information from published academic research studies, researchers can now develop better Nrf2 dietary supplements.
As of 2007,�research studies have demonstrated the complex function of the Nrf2 pathway. Nrf2 activators have been found to mimic factors of different structures within the human body. Through these pathways, Nrf2 activators have been equipped to feel changing conditions throughout the cell in order to keep balance and respond to the evolving requirements of the genes.
Why Use Nrf2-Activating Supplements?
As Nrf2-activation abilities diminish with age in organisms, changes may begin to occur. Research studies have demonstrated that the focus of Nrf2 in cells declines with age, showing increased markers of oxidative stress. A variety of age-related diseases like atherosclerosis and cardiovascular disease, arthritis, cancer, obesity, type 2 diabetes, hypertension, cataracts, and Alzheimer’s disease as well as Parkinson’s diseases can develop due to these changes. Oxidative stress has been found with these health issues.
By stimulating the cell’s capacity to increase the production of Nrf2 activators, Nrf2 dietary supplements can help revive the human body’s own ability to counteract the effects of oxidative stress. Polyunsaturated fatty acids, or PUFAs, are one of the most readily oxidized molecules and they’re particularly vulnerable to suffer damage from free radicals. Thiobarbituric acid, or TBARS, production can increase with age, indicating heightened oxidative stress along with a drop in Nrf2-regulated pathways.
Biologically, gene induction is a really slow mechanism, generally requiring hours to transfer through a pathway. As a result,�many enzymes possess their very own on/off switches which could be triggered in minutes by different regulatory enzymes. Researchers have developed proprietary compositions of Nrf2 activators which utilize this knowledge base of activation. Nrf2 activation is composed not just of the Nrf2 transcription factor being discharged from its inhibitor and migrating to the cell nucleus, but also binding to specific DNA sequences to encourage cytoprotective gene expression, regulating the pace at that Nrf2 is taken out of the nucleus.
Understanding the elimination procedure and the activation of Nrf2 in the human body has allowed researchers to build combinations of different Nrf2 activators to accomplish the reflection of genes through its modulation. The combination of the knowledge base, together with the wide variety of other research studies has�helped produce Nrf2 activators for use as dietary supplements. The scope of our information is limited to chiropractic and spinal health issues. To discuss the subject matter, please feel free to ask Dr. Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topic Discussion: Relieving Knee Pain without Surgery
Knee pain is a well-known symptom which can occur due to a variety of knee injuries and/or conditions, including�sports injuries. The knee is one of the most complex joints in the human body as it is made-up of the intersection of four bones, four ligaments, various tendons, two menisci, and cartilage. According to the American Academy of Family Physicians, the most common causes of knee pain include patellar subluxation, patellar tendinitis or jumper’s knee, and Osgood-Schlatter disease. Although knee pain is most likely to occur in people over 60 years old, knee pain can also occur in children and adolescents. Knee pain can be treated at home following the RICE methods, however, severe knee injuries may require immediate medical attention, including chiropractic care.
How important is nutrition for our brain health? In the current work force, we are continuously stressed, often forced to finish tasks faster in order to meet ever so demanding deadlines. In addition, we are expected to maintain our optimal mental health, as this can be an essential�part towards delivering quality work. When our mental health is being affected by our hectic lifestyles, however, several practices which can help you start thinking more clearly can include sleeping properly, controlling stress, and even taking nutritional supplements for your brain health.
One nutritional supplement which has been widely recognized for its ability to boost brain health is curcumin, the active ingredient found in turmeric. Well-known for its antioxidant properties and its capacity to control inflammation in the human body, this powerful herb can also promote good mood and cognition. Another specific group which has reported significant benefits with the increased use of curcumin, is the elderly population. Below, we will discuss how curcumin can help boost brain health as well as demonstrate additional benefits this golden gem can have on our overall health and wellness.
Curcumin: a Golden Gem for Brain Health
In accordance with the Journal of Pharmacology, curcumin is made-up of a variety of substances which can encourage biological mechanisms that counteract age-related cognitive decline, dementia, or mood disorders. One randomized, double-blind, placebo-controlled trial analyzed the acute, of approximately 1 and 3 hours following a single dose, chronic, of approximately 4 weeks, and acute-on-chronic, of approximately 1 and 3 hours after one dose subsequently after chronic treatment, consequences of a curcumin formulation on cognitive function, mood, and blood biomarkers in 60 healthy adults ranging from the 60 to 85 years of age. After about one hour of application, the curcumin had considerably enhanced the participant’s functionality on attention and working memory tasks, in comparison with the placebo. Working memory and mood, which included general fatigue, change in calmness, contentedness and fatigue triggered by emotional strain were fundamentally improved following chronic therapy.
Curcumin boosts BDNF (brain-derived neurotrophic factor), the brain hormone which helps boost the development of new neurons that are in charge of improving memory and learning as well as supplying a substantial option for countering the aging brain. Additionally, this powerful ingredient increases blood circulation to the brain, also providing a much better attention span for greater work productivity.
Appreciating its anxiolytic effects can be one of the greatest benefits of carrying curcumin. According to the Journal of Clinical Psychopharmacology, a randomized double-blinded and double-blind trial with 60 subjects experiencing stress-related symptoms, including exhaustion, were to get routine curcumin nutritional supplements, and placebo for 30 days. The results indicated a greater quality of life, and diminished stress and fatigue for those receiving regular curcumin intakes. This progressive compound is believed to be able to help alleviate depression by altering the release of dopamine and serotonin, two powerful hormones which help keep the human mind and body at ease. Curcumin also promotes the optima health and wellness of inflammation pathways from the brain, which ultimately will help improve energy, mood, and production levels.
Curcumin may additionally promote cognition via its powerful antioxidant action which improves the bioavailability of DHA, the potent omega-3 fatty acid demonstrated to boost brain health. A research study in the American Journal of Geriatric Psychiatry revealed that curcumin really does protect the brain from neurodegeneration. The evaluation and analysis included 40 participants ranging from the ages of 51 to 84 years of age. Each individual subject consumed 90mg of curcumin twice per day or placebo for 18 weeks. The results indicated enhanced long-term healing, visual memory, and focus. With its tremendous medicinal properties, curcumin can also support neuroplasticity, which empowers the brain to change and fortify itself even through the natural degeneration with aging.
Curcumin can also promote anti-seizure action. With its antioxidant properties, this golden gem can help slow down reactive astrocyte expression, which helps cells survive within the mind. According to the Neuropharmacology Laboratory, Department of Pharmacology, the antioxidant properties of curcumin helped alleviate migraines, cognitive impairment, and cognitive stress in rats. A dental pre-treatment of curcumin was given to male rats which were additionally treated together with Pentylenetrazole, or PZT, every other day. The study demonstrated that curcumin enhanced the seizure score and indicated a diminished amount of myoclonic jerks. Furthermore, the outcome measures of the research study demonstrated that curcumin restructures seizures, oxidative stress, and brain function. Moreover, it helps protect memory function which may also be jeopardized by seizure activity.
Using its capability to strengthen fatty acids in the mind, curcumin helps athletes achieve better physical performance by boosting critical thinking, improving problem solving, and developing improved choices. The neuroprotective properties in curcumin also help regenerate tissues. In reality, based on Stem Cell Research and Therapy, a research study was conducted between the effects of curcumin on endogenous stem cells which were impartial. The study demonstrated that curcumin played an essential role in the healing of cells from combating the activation of microglia cells. Scientists in the Institute of Neuroscience and Medicine in Julich, Germany, observed the effects of impartial stem cell generation. During a 72-hour period, the evaluation and analysis demonstrated and indicated that the turmeric curcumin improved cellular generation by up to 80 percent. This shows how powerful curcumin could be for successful brain health function.
Dr. Alex Jimenez’s Insight
Nutrition is a fundamental factor in overall health and wellness. In today’s stressful world, however, it can often become difficult to eat a proper meal, let alone making sure we are taking in all the necessary nutrients we require on a regular basis. That, plus the added pressure of the workforce can have detrimental effects on our brain health. Dietary supplements, such as curcumin, have been demonstrated to have tremendous benefits on brain health. Although we may not always have the “free time” to sit down and have a properly balanced meal, taking nutritional supplements like curcumin, among others, can help improve the human body’s general well-being.
While many research studies have found that natural remedies and botanicals, such as dietary supplements apart from vitamins and minerals, continue to be the most common complementary health approach in the United States today, more and more alternative treatment options, such as chiropractic care, have started to incorporate these into their practices. As a matter of fact, a majority of chiropractors give nutritional advice, as well as recommendations for other lifestyle recommendations, as a general part of their treatment plan. Because chiropractic care is based on the notion of naturally treating the human body as a whole, enhancing it’s own healing properties without the use of drugs and/or medications as well as other invasive procedures, this healthcare profession relies on offering the necessary health maintenance components for optimal health and wellness. These components can include nutrition, water, rest, exercise, and clean air. Many chiropractors also offer curcumin supplements to help promote recovery.
This exceptional nutritional supplement, curcumin, helps improve mental clarity, improve cognition, improve endurance, and supplies anxiolytic benefits. Whether it’s more work fabricating, or a much better disposition, curcumin is a hidden golden gem for health.�The scope of our information is limited to chiropractic as well as to spinal injuries and conditions. To discuss the subject matter, please feel free to ask Dr. Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topics: Back Pain
Back pain is one of the most prevalent causes for disability and missed days at work worldwide. As a matter of fact, back pain has been attributed as the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience some type of back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments and muscles, among other soft tissues. Because of this, injuries and/or aggravated conditions, such as herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief.
Allergy Sufferers!�As winter gives way to spring, seasonal allergies can really get you down. Whether you get a few sniffles and some sneezing or you are down for the count with every terrible allergy symptom known to man, it can make spring pretty unbearable.
There is no shortage of allergy medications on the market, but they come with their own issues. The majority of them cause drowsiness and other unpleasant side effects, leaving you barely able to function. Those that are made from a �non drowsy� formula sound great, but if you have certain health conditions, like high blood pressure, you are out of luck � and stuck either taking the ones that make you sleep sucking it up and dealing with your allergies sans medication.
That�s no way to live.
What Are Allergies?
When your immune produces histamines in response to an allergen that you encounter the physiological reaction that you experience is broadly referred to as allergies or hay fever. The allergens may be simple substances that normally do not affect people, but when your body is out of balance, it can cause a variety of problems.
Symptoms of allergies include:
Runny nose
Stuffy nose
Headache
Sneezing
Itchy eyes
Coughing or scratchy throat
Skin Rash or Hives
Swelling
Diarrhea
Nausea
Fatigue
Anaphylaxis, severe, life threatening allergies can include swelling of the airways, tongue, and throat, inability to breathe due to blocked airway, and other dangerous symptoms.
The allergens can be something you come in contact with, like poison ivy, something you breathe in, like mold or dust, or it can be something you ingest, like strawberries or peanuts. Different people will have different allergies, but those who are allergic to the same things may not have the same reaction. Often a doctor or allergist will diagnose your allergies.
Chiropractic Care For Allergy Sufferers
Chiropractic treatments have been found to be very effective for relieving allergy symptoms and even stopping allergies at their source. It reduces the severity of allergy symptoms as well as the frequency of occurrence. It does not work like allergy medications which have an anti-histamine effect and only work as a short term fix for your allergy symptoms.
Chiropractic treatments help your body become more balanced so that it is better equipped for combating allergies at the source. When your spine is not aligned it can impact your nervous system leading to a variety of problems � including allergies. Your immune system can be affected, causing it to malfunction.
A chiropractor can help relieve the stress on your nervous system by aligning your spine. This takes the pressure off of nerves, allowing your immune system to function at a more optimal level. This makes it easier for your body to ward off infections while recognizing allergens as harmless.
When your immune system encounters allergens it doesn�t overreact to them. Instead, the reaction is much more subdued, or even nonexistent. Chiropractic has also been found to help asthma patients breathe easier. Asthma symptoms are diminished.
Chiropractic care is more than just spinal manipulation, though. It promotes whole body wellness. Patients are taught exercise, stress relief, and nutrition so that the entire system is treated. The whole body treatment plan for chiropractic patients will help you be allergy free in a short time.
It is important to follow your chiropractic plan thoroughly and consistently. Get plenty of rest and take time to destress. The more you can relax and take care of yourself, the healthier you will be overall. Chiropractic care can help so many health conditions; it can actually make you healthier. Allergy sufferers or if you are struggling with allergies for the first time, give chiropractic care a try you just might be surprised.
IFM's Find A Practitioner tool is the largest referral network in Functional Medicine, created to help patients locate Functional Medicine practitioners anywhere in the world. IFM Certified Practitioners are listed first in the search results, given their extensive education in Functional Medicine













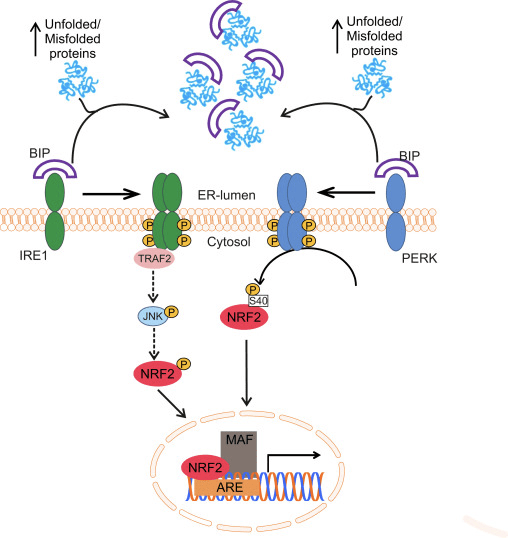
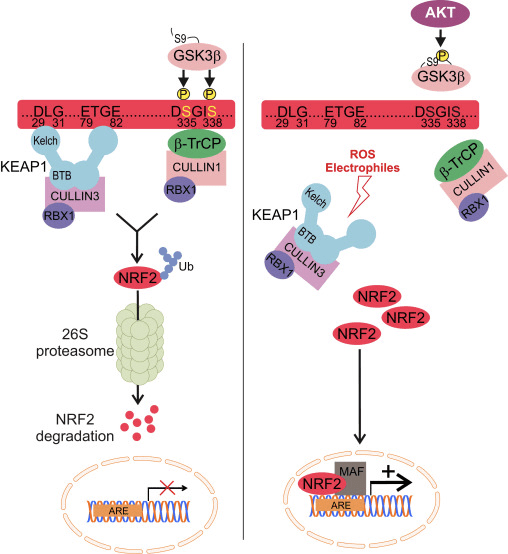
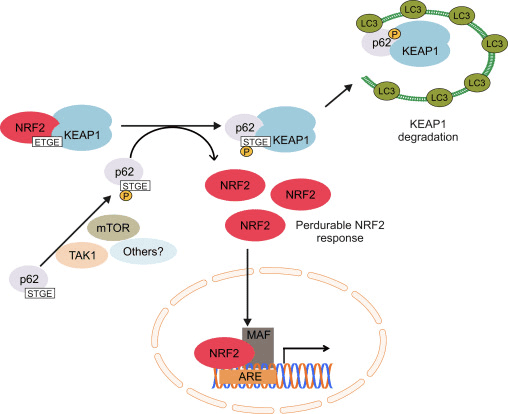
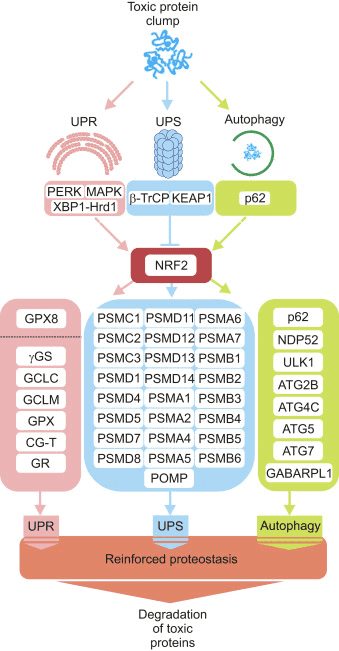







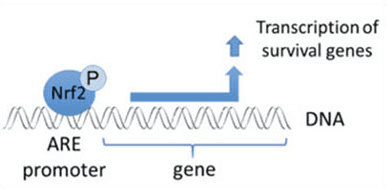
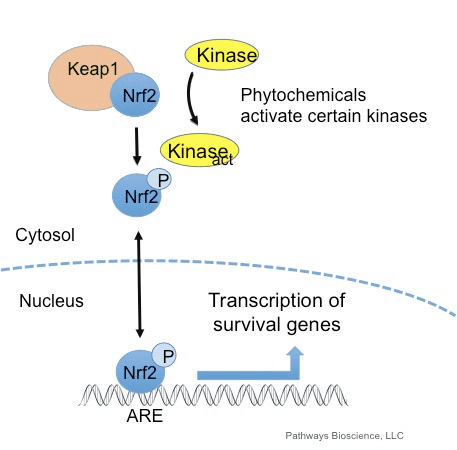





 Chiropractic Care For Allergy Sufferers
Chiropractic Care For Allergy Sufferers





