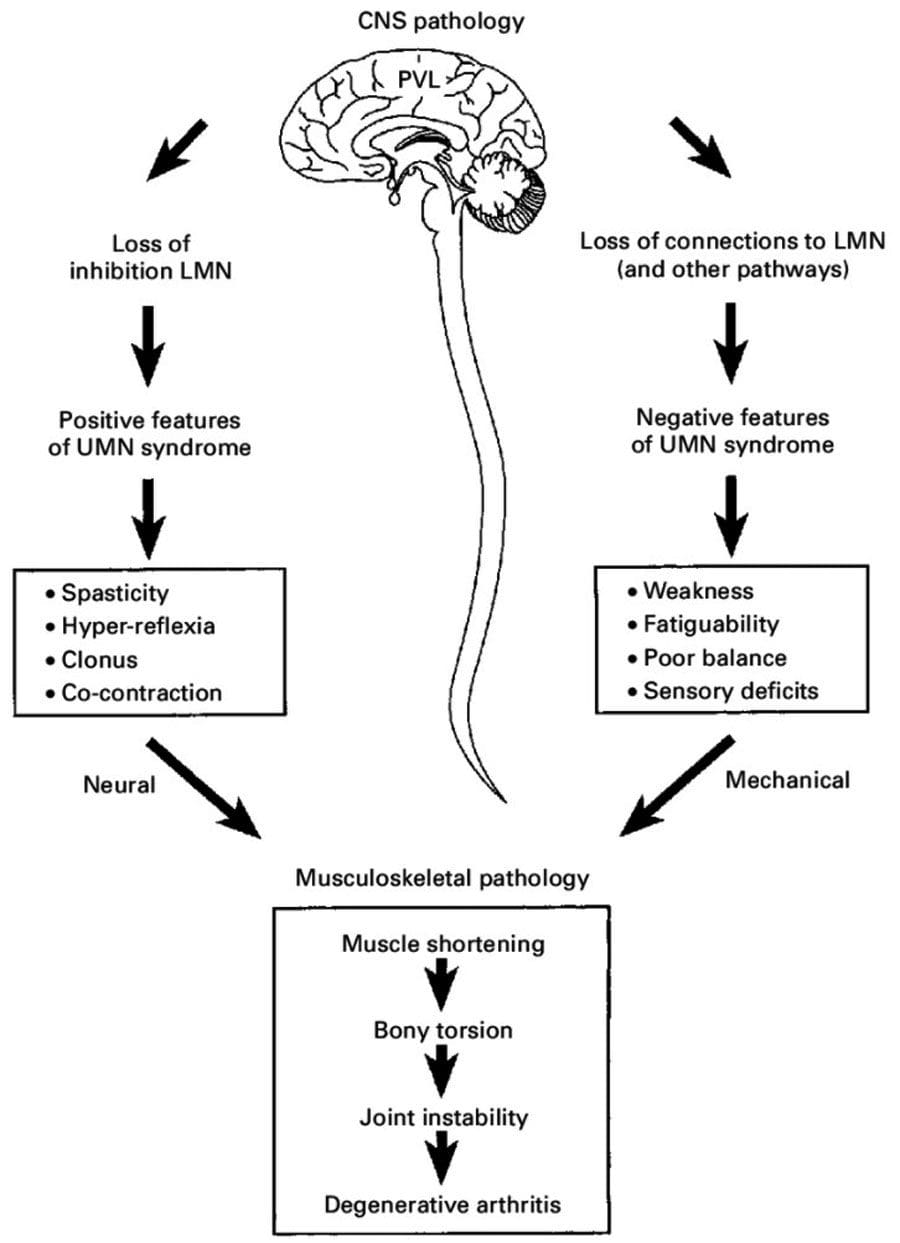
Chiropractic is a form of neuromusculoskeletal care that repairs and re-optimizes the nervous system that helps to relieve tension, inflammation, pain and restore body health. Like any piece of machinery that requires regular maintenance to operate correctly without issues, so too is the body an exquisite machine that needs regular maintenance. Treatment restores and maintains optimal function to the nervous system, muscles, bones, tendons, ligaments, and joints.
Neuromusculoskeletal Care
Central Nervous System – CNS
The Central Nervous System or CNS is the body’s computer.
It processes functions in the body and mind.
It s made up of the brain and spinal cord.
It helps process external information through sight, sound, touch, smell, and taste.
The CNS interprets sensory information that goes through the spinal cord to the brain, and the body responds accordingly.
It controls voluntary movements like walking, talking, exercising.
It controls involuntary movements like blinking, breathing, digestion.
It allows thoughts, emotions, and perceptions to be created and expressed.
When the Central Nervous System works correctly, the body is coordinated, all senses send and receive information effectively, and cognition/thinking is clear.
Peripheral Nervous System – PNS
The Peripheral Nervous System or PNS transmits signals from the outside to the Central Nervous System.
The PNS is comprised of all the nerve bundles that run throughout the body.
The PNS is responsible for transmitting information back and forth from the CNS to the rest of the body by electrical impulses and current.
When the PNS is functioning correctly, and there is thorough nerve circulation throughout the body, there are no issues like numbness, weakness, pain, and digestion is in top form.
Enteric Nervous System – ENS
The Enteric Nervous System or ENS is a branch of the peripheral nervous system that signals the digestive system.
It comprises nerves that stretch across the entire gastrointestinal tract.
It allows the nervous system to communicate with the digestive tract and regulate activity.
It can stop the digestive process when the body is under stress or strain, i.e., fight or flight response.
Inflammation and stress can cause the enteric system to disrupt and cause digestive problems.
Pain Relief
Body pain can be an indication of an imbalance in the nervous system. Misalignment from work, home activities, trauma, injury, or posture problems, a subluxation/misalignment can turn into discomfort that leads to pain and neuromusculoskeletal system issues. Pain in the neuromusculoskeletal system can result from spinal misalignment and a shifting skeletal structure. The joints and misaligned bones compress the surrounding nerves and tissue, causing inflammation that results in pain symptoms. This causes the muscular system to compensate for the lack of stability support from the skeletal structure causing awkward body posturing. A chiropractor realigns the spinal structure, removing the compression/pressure from the nerves, tissues, muscles, ligaments, and tendons.
Body Composition
Chiropractic Medicine
The nervous system is highly responsive to chiropractic care because of chiropractic’s focus on the spine, which restores and rejuvenates. Chiropractic medicine and a properly aligned spine:
Reduces and eliminates pain.
Reduces and eliminates headaches and migraines.
Improves balance and coordination.
Improves digestive function.
Improves quality of sleep.
Increases energy.
Increases flexibility and mobility.
Enhances cognition and clear thinking.
References
Goudman, Lisa et al. “The Link Between Spinal Cord Stimulation and the Parasympathetic Nervous System in Patients With Failed Back Surgery Syndrome.” Neuromodulation: journal of the International Neuromodulation Society vol. 25,1 (2022): 128-136. doi:10.1111/ner.13400
Gyer, Giles et al. “Spinal manipulation therapy: Is it all about the brain? A current review of the neurophysiological effects of manipulation.” Journal of integrative medicine vol. 17,5 (2019): 328-337. doi:10.1016/j.joim.2019.05.004
Millet, Guillaume Y et al. “The role of the nervous system in neuromuscular fatigue induced by ultra-endurance exercise.” Applied physiology, nutrition, and metabolism = Physiologie appliquee, nutrition et metabolisme vol. 43,11 (2018): 1151-1157. doi:10.1139/apnm-2018-0161
Stoll, T et al. “Physiotherapie bei lumbaler Diskushernie” [Physiotherapy in lumbar disc herniation ]. Therapeutische Umschau. Revue therapeutique vol. 58,8 (2001): 487-92. doi:10.1024/0040-5930.58.8.487
Chiropractors understand the need to treat the whole body to reduce and alleviate pain. Many types of pain are caused by inflammation. Inflammation is a natural and healthy response to injury; however, chronic inflammation is not. Low-grade chronic inflammation can lead to health problems like obesity, diabetes, arthritis, dementia, heart disease, stroke, and cancer. If not treated, chronic inflammation can spread throughout the body, causing pain and aggravation. When it comes to nutritional health, the foods can worsen chronic pain. Chiropractors and doctors recommend an anti-inflammation diet for chronic pain.
Anti-Inflammation Diet
When the body gets injured, the nervous system sends signals to the immune system to send chemicals and new red and white blood cells needed for healing. The immune system works correctly when it fights infection by activating when the body recognizes anything foreign entering the system. This could be plant pollen, chemicals, or invading microbes. Studies show that individuals get 50 percent of their calories from sugar, white flour, vegetable oil, and industrial seed oils. These foods are known to increase inflammation in the body. Eating foods that trigger the immune system, like refined white flours or sugar, creates inflammation that does not turn off because the information sent constantly signals an injury exacerbating inflammation and pain.
Foods That Cause Inflammation
The following foods should be avoided or limited as much as possible:
Sugary beverages and soda drinks.
Margarine and lard.
Gluten and white pasta.
Refined carbohydrates in white bread and pastries.
Processed meat like sausages and hot dogs.
Red meat like steaks and burgers.
Fried foods that are high in trans fats, like chips and fries.
Excessive alcohol.
Some of these foods have been associated with chronic diseases that include:
Foods high in natural antioxidants include apples and blueberries. Antioxidants are reactive molecules that can reduce the number of free radicals in the diet. A free radical is a molecule that has the potential to alter and damage the cells in the body. Damaged cells increase the risk of disease development.
Chiropractic Inflammation Relief
Chiropractic physiotherapy strengthens the body by strengthening the immune system removing any blockage. This maintains the natural flow of blood and nutrients throughout the body, increasing the immune system’s defense. Changing to an anti-inflammation diet can help boost the immune system but can be a challenging adjustment. Discuss available options to manage pain relief and inflammation.
Body Composition
Muscle Is Not Just For Athletes
Many individuals think that muscle gain is only necessary for athletes. Not everyone wants to be muscular, but everyone needs to be able to fight off sickness from infection/s. Muscle is made up primarily of water and protein. Protein is an essential macronutrient that the body needs to function correctly. When the body enters a stressed state like becoming sick, the body’s protein demands suddenly increase up to four times the amount usually required. If the body does not get the necessary protein from the food, it will begin to take what it needs from the muscles and start breaking them down. If muscles aren’t sufficiently developed or underdeveloped, the body becomes reduced in its ability and strength to fight off infections and increases the chances for future ones.
References
Haß, Ulrike et al. “Anti-Inflammatory Diets and Fatigue.” Nutrients vol. 11,10 2315. 30 Sep. 2019, doi:10.3390/nu11102315
Owczarek, Danuta et al. “Diet and nutritional factors in inflammatory bowel diseases.” World journal of gastroenterology vol. 22,3 (2016): 895-905. doi:10.3748/wjg.v22.i3.895
Sears, Barry. “Anti-inflammatory Diets.” Journal of the American College of Nutrition vol. 34 Suppl 1 (2015): 14-21. doi:10.1080/07315724.2015.1080105
After all of these years, I am happy to announce that the Texas Supreme Court has finally made a decision regarding the Texas Board of Chiropractic Examiners et al v. Texas Medical Association case on January 29th, 2021. With great honor and gratitude, I’d like to continue to extend sincere thanks to everyone who worked hard on this case and whose tremendous efforts resulted in the decision. Thanks to the Supreme Court’s decision, chiropractors in Texas can now carry on their jobs accordingly. Below, I have provided a letter from Board President, Mark R. Bronson, D.C., F.I.A.N.M. on behalf of the Texas Board of Chiropractic Examiners stating the Texas Supreme Court’s decision in the Texas Board of Chiropractic Examiners et al v. Texas Medical Association case on January 29th, 2021. – Dr. Alex Jimenez D.C., C.C.S.T.
February 1, 2021
On behalf of the Texas Board of Chiropractic Examiners, I extend our sincere thanks and appreciation to everyone whose efforts resulted in the Texas Supreme Court’s decision in Texas Board of Chiropractic Examiners et al v. Texas Medical Association on January 29, 2021. Special thanks are due to all the attorneys at the Office of the Attorney General who worked on this case over these years.
The decision properly affirmed the validity of the Board’s scope of practice rule, which the court clearly said does not exceed our statutory scope of chiropractic practice. The court unequivocally held that the Board�s rules do not violate Occupations Code Chapter 201 or run counter to the chapter’s objectives set by the Texas Legislature, and in fact, carefully observe the statutory boundary between the medical and chiropractic professions. This decision, which recognizes the common sense and long-standing inclusion of associated nerves in chiropractic diagnosis and treatment, preserves and strengthens the essence of chiropractic.
Thanks to the court’s decision, our licensees can now fulfill their duties as vital portal-of-entry healthcare providers in Texas without fear. The court’s decision reaffirms the principles of economic freedom that have made Texas the best state in the nation to be a chiropractor.
Sincerely,
Mark R. Bronson, D.C., F.I.A.N.M. Board President
Texas Board of Chiropractic Examiners
The scope of our information is limited to chiropractic, musculoskeletal, physical medicines, wellness, and sensitive health issues and/or functional medicine articles, topics, and discussions. We use functional health & wellness protocols to treat and support care for injuries or disorders of the musculoskeletal system. Our posts, topics, subjects, and insights cover clinical matters, issues, and topics that relate and support directly or indirectly our clinical scope of practice.*
Our office has made a reasonable attempt to provide supportive citations and has identified the relevant research study or studies supporting our posts. We also make copies of supporting research studies available to the board and or the public upon request. We understand that we cover matters that require an additional explanation as to how it may assist in a particular care plan or treatment protocol; therefore, to further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900. The provider(s) Licensed in Texas & New Mexico*
PODCAST: Dr. Alex Jimenez, Kenna Vaughn, Lizette Ortiz, and Daniel “Danny” Alvarado discuss nutrition and fitness during these times. During quarantine, people have become more interested in improving their overall health and wellness by following a proper diet and participating in exercise. The panel of experts in the following podcast offers a variety of tips and tricks on how you can improve your well-being. Moreover, Lizette Ortiz and Danny Alvarado discuss how they’ve been helping their clients achieve their optimal well-being during these COVID times. From eating fruits, vegetables, lean meats, good fats, and complex carbohydrates to avoiding sugars and simple carbohydrates like white pasta and bread, following a proper diet and participating in exercise and physical activity is a great way to continue to promote your overall health and wellness. – Podcast Insight
If you have enjoyed this video and/or we have helped you in any way
please feel free to subscribe and share us.
Thank You & God Bless.
Dr. Alex Jimenez RN, DC, MSACP, CCST
PODCAST: Dr. Alex Jimenez and Dr. Marius Ruja discuss the importance of personalized medicine genetics and micronutrients for overall health and wellness. Following a proper diet and participating in exercise alone isn’t enough to make sure that the human body is functioning properly, especially in the case of athletes. Fortunately, there are a variety of tests available that can help people determine if they have any nutritional deficiencies that may be affecting their cells and tissues. Vitamin and mineral supplements can also ultimately help improve an individual’s overall health and wellness. While we may not be able to change certain aspects of our genes, Dr. Alex Jimenez and Dr. Marius Ruja discuss that following a proper diet and participating in exercise while taking the proper supplements, can benefit our genes and promote well-being. – Podcast Insight
If you have enjoyed this video and/or we have helped you in any way
please feel free to subscribe and share us.
Thank You & God Bless.
Dr. Alex Jimenez RN, DC, MSACP, CCST
IFM's Find A Practitioner tool is the largest referral network in Functional Medicine, created to help patients locate Functional Medicine practitioners anywhere in the world. IFM Certified Practitioners are listed first in the search results, given their extensive education in Functional Medicine