People will often visit the doctor looking for hip pain relief while other people will often attempt to take care of it themselves. Hip pain can frequently occur due to an injury or underlying condition as well as a variety of health issues. However, not many people who visit the doctor or attempt to take care of their hip pain expect it to originate as a result of sciatica. Many people’s misunderstanding concerning the origin of their pain is due to not understanding back and hip anatomy. �
Contents
Understanding Back Pain
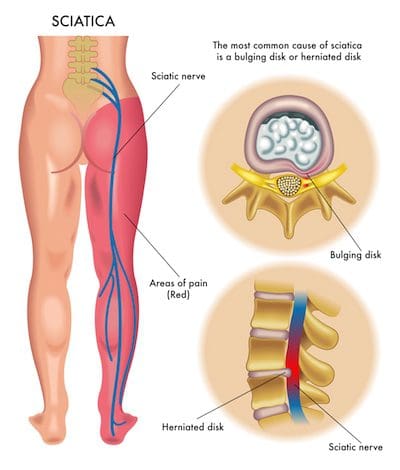
Because of the complexity of the spinal cord and its nerve roots, back pain can often radiate or travel down buttocks, hips, and thighs, sometimes even extending as far down into the legs, knees, and feet.� Sciatica, or sciatic nerve pain, is a collection of symptoms, rather than a single injury or underlying condition, caused by the compression or impingement of the sciatic nerve. Sciatica is characterized by pain, tingling sensations, and numbness along the length of the sciatic nerve. �
The sciatic nerve is the largest and longest nerve in the human body. It extends from the lower back, down the buttocks, hips, and thighs, into the legs, knees, and feet. The irritation or inflammation of the sciatic nerve may also cause painful symptoms in the upper extremities. It’s essential to understand that an injury or underlying condition, as well as a variety of health issues, can cause back pain and sciatica. Sciatica is generally a symptom associated with numerous other medical problems. �
By way of instance, a herniated disc can cause back pain and sciatica. People may experience “hip pain”, however, the painful symptoms may not necessarily be indicative of a hip joint health issue. Degenerative disc disease can cause a narrowing of the spinal canal, referred to as spinal stenosis, which may cause hip pain. Spinal stenosis can cause hip pain with physical activity and fatigue. Spinal stenosis symptoms are relieved with sitting and will re-occur when physical activity is resumed. �
You will find differences in painful symptoms involving a herniated disc, spinal stenosis pain, and other health issues. Painful symptoms associated with a herniated disc often worsen when sitting and improve with physical activity. A herniated disc, much like spinal stenosis, can also cause sciatica and it may often be due to degenerative disc disease. Sciatica, or sciatic nerve pain, will generally radiate or travel down the buttocks, hips, and thighs into the legs, knees, and feet. �
Understanding Hip Pain
If the health issue originates in the hip joint itself, common painful symptoms may often include groin pain on the affected side, which can occasionally radiate down the inner region of the thigh into the front of the leg. The painful symptoms can also travel to the knee, making healthcare professionals and patients believe their pain is associated with a knee problem rather than a hip problem. Walking worsens the pain and with continued physical activity, the pain increases. �
Hip pain caused by health issues, such as arthritis, can cause painful symptoms to gradually worsen over time.� Minimal physical activity, even slight movements while in bed, can also ultimately worsen the painful symptoms. Other health issues, such as advanced congenital hip dysplasia or avascular necrosis of the hip, can also cause these painful symptoms. When the hip pain originates due to arthritis, the movements of the hip joint may often be limited or restricted. �
Occasionally, hip pain can be a result of another health issue referred to as bursitis. A weak abductor muscle, a leg length discrepancy, overuse, and a deteriorating hip joint can ultimately cause bursitis. Often times, the true source of a patient’s bursitis cannot be determined. Painful symptoms associated with bursitis include pain and discomfort on the side of the hip with prolonged walking, lying sideways in bed or when rising or standing from a chair as well as with similar movements. �
Sciatica Diagnosis
A comprehensive medical history and physical evaluation can help healthcare professionals determine the source of the patient’s symptoms. X-rays demonstrate specific bony/cartilage modifications, however, x-rays don’t always necessarily demonstrate health issues associated with soft tissues, such as tendons, ligaments, muscles, and even nerves. X-rays may also help diagnose degenerative disc disease when the patient is feeling well and not experiencing any painful symptoms. �
As previously mentioned above, because x-rays only demonstrate certain bony/cartilage changes, it’s essential for healthcare professionals to also request the patient to have an MRI, or magnetic resonance imaging, to confirm the presence of health issues associated with soft tissues, such as tendons, ligaments, muscles and nerve roots. That’s why it’s essential to undergo the background and physical evaluation which could confirm the diagnosis before beginning with the best treatment. �
Sciatica Treatment
The two types of health issues are frequently treated utilizing anti-inflammatory drugs and/or medications, especially in mild to moderate instances. Some types of analgesics can also be utilized intermittently. It is essential to understand that both health issues can be improved considerably through weight loss, stretches and exercises. Epidural blocks will help several types of spine health issues. Utilizing a cane when walking might help improve back pain, hip pain, and sciatica. �
Surgical interventions, whether if it’s a hip replacement for hip arthritis or spine surgery due to a herniated disc, spinal stenosis, or any other type of health issue, is generally considered to be the last resort for treating back pain, hip pain, and sciatica. Full physical evaluations are recommended and conservative treatment approaches are often tried first. Remember, there are many treatment options for hip pain and sciatica. Diagnosis is the first step to effective treatment. �
Sciatica, or sciatic nerve pain, is a collection of symptoms, rather than a single injury or condition, characterized by pain and discomfort, tingling sensations, and numbness anywhere along the length of the sciatic nerve. The sciatic nerve is the longest and largest nerve in the human body which extends from the lower back, down the buttocks, hips, and thighs, into the legs, knees, and feet. The compression or impingement of the sciatic nerve and irritation due to a herniated disc, among other health issues, can ultimately cause sciatica symptoms and low back pain. – Dr. Alex Jimenez D.C., C.C.S.T. Insight
Low Back Pain
The purpose of the article was to discuss and demonstrate hip pain associated with sciatica. Sciatica is a collection of symptoms characterized by pain and discomfort, tingling sensation, and numbness. The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues as well as functional medicine articles, topics, and discussions. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 . �
Curated by Dr. Alex Jimenez �
Additional Topic Discussion: Foot Orthotics
Low back pain and sciatica are common health issues which affect many individuals worldwide. However, did you know that chronic pain may be due to foot problems? Health issues originating in the foot may ultimately cause imbalances in the spine, such as poor posture, which can cause the well-known symptoms of low back pain and sciatica. Custom foot orthotics, individually designed with 3-arch support can help promote overall health and wellness by supporting and promoting good posture and correcting foot problems. Custom foot orthotics can ultimately help improve low back pain and sciatica. �
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
�
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download �
* All of the above XYMOGEN policies remain strictly in force. �
Asking questions is the best way to learn, of course, and chiropractic patients often ask questions about some of the health issues they face. Some chiropractic patients are curious about back problems, for example, because chiropractors are well known for being extremely knowledgeable when it comes to spine health and the musculoskeletal system. A few chiropractic patients want to know about TMJ disorders.
Here are a few of the most common things chiropractic patients want to know about TMJ disorders.
Contents
The Most Frequently Asked Questions about TMJ Disorders
�TMJ disorder?
A TMJ disorder affects the temporomandibular joint (TMJ), which is the joint where the jawbone connects to the head, just in front of the ear. The bones and muscles of the TMJ allow the jaw to move up and down, and side to side, enabling a person to talk, chew and yawn.
TMJ disorders (TMDs) can cause pain in the jaw joint and in the muscles that control the movement of the jaw.
How common is it?
About 12 percent of the population experiences symptoms of TMJ at any given time, according to the TMJ Association.
The Causes?
There are several known causes of TMJ disorders, including:
Dental procedures, especially those that require the mouth be open for a long time
Insertion of a breathing tube prior to surgery
Arthritis
Misalignments of the jaw
Grinding teeth, known as bruxism, may cause TMJ disorders but not everyone who grinds their teeth develops a TMD.
Medical professionals may not be able to determine the underlying cause of TMJ disorders in many cases but chiropractors can treat TMJ disorders even if the cause cannot be determined.
The Symptoms of TMJ disorder?
Many people with TMD describe their symptoms as a dull, aching pain that comes and goes in their jaw joint and in nearby areas. Some people with TMJ disorders do not experience pain but have trouble moving their jaws.
Other common symptoms of TMJ disorders include:
Pain or stiffness in the jaw muscles
Chronic headaches
Neck, shoulder pain
Clicking, popping or grating of the jaw joint when the individual opens or closes his or her mouth
Limited movement of the jaw
�Locking� of the jaw
Tinnitus, which includes ear pain, fullness, pressure, and ringing in the ears
Dizziness, vision problems
A bite that feels �off� when the person closes his or her mouth
Treatment for TMJ disorders?
The best way to treat TMJ disorders is to eat soft foods, ice the affected area with ice packs to ease pain and try to avoid extreme jaw movements, such as chewing gum or yawning.
Does Chiropractic treat TMJ disorders?
Yes, chiropractic safely and effective treats TMJ disorders. A chiropractor will perform a full evaluation of the patient�s temporomandibular joint and surrounding bones, muscles, tendons and ligaments to assess the full scope of the TMJ problem and determine its underlying causes. The chiropractor can then suggest a range of treatment that can include stretches and exercises that alleviate pain and melt stiffness in the jaw joint. The chiropractor can also detect and address any misalignments that may be contributing to TMJ pain.
For more information about the temporomandibular joint, TMJ disorders and chiropractic for TMJ problems, contact a chiropractor with experience in diagnosing and treating TMJ problems.
Reduce *IMBALANCE & PAIN* with Functional Orthotics | El Paso, Tx
Foot pronation is a natural movement during standing, walking, or running, however, excessive foot pronation can cause postural imbalances which can result in chronic pain, including low back pain and sciatica. Dr. Alex Jimenez, a chiropractor in El Paso, TX, can help diagnose and treat a variety of health issues associated with foot problems through the utilization of functional custom foot orthotics.
Functional custom foot orthotics are specially designed to accommodate every person’s unique foot anatomy. Excessive foot pronation can ultimately lead to poor posture, which can add unnecessary pressure to the spine and it’s surrounding structures. Dr. Alex Jimenez is the non-surgical choice for foot problems and other health issues through the use of functional custom foot orthotics.
Custom Orthotics & Store Bought Insoles
Store-bought orthotic insoles are cheap but the cost to you and your health can be very high. Understanding the differences between over the counter and custom made orthotics is important. Foot Levelers customized orthotics provide the highest-quality and most effective orthotic out there.
Foot Levelers custom orthotics are individually designed for your feet. This achieves a balanced foundation and a stabilized pelvis. These orthotics are handmade based on 3D scans or casts, and your doctor�s examination.
Over-the-counter insoles do cost less, but they only support one arch in the foot. When only one arch is supported, the structure can collapse, and that’s when problems can begin in other parts of the body. Over-support of one arch can cause pain and symptoms, instead of relieving them.
Orthotics Treat Way More Than Feet
Radiculopathy is associated with some of the most significant causes of chronic or acute low back pain. However, it is important to note that the condition itself does not cause pain. Instead, elements of radiculopathy, such as disc herniation, nerve root impingement, and facet arthropathy are actually what causes the pain.
Radiculopathy is a condition of the spine that occurs when a nerve is compressed, causing pain, weakness, tingling, or numbness along the nerve�s course. In the lower back, that course is in the leg. While it is most common in the lower back, radiculopathy can occur in the cervical or thoracic regions of the spine as well.
NCBI Resources
The�temporomandibular joints, TMJ, are the lower jaw hinges that sit on either side of the head in front of each ear. They are responsible for the lower jaw opening, closing, sliding, and rotating. The TMJs are the most body�s most complex joints. The typical person uses them more than 5,000 times a day by talking, laughing, yawning, chewing, eating, smiling, and swallowing.
Sciatica is commonly associated with low back pain, however, sciatica may also generally occur due to a variety of other health issues. The sciatic nerve is the longest and largest nerve in the human body. The sciatic nerve extends from the lower back, or lumbar spine, down the buttocks, hips, and thighs, into the legs, knees, and feet. The sciatic nerve is in charge of controlling many muscles in the legs and it also provides feeling to the skin of the majority of the lower extremities. �
Sciatica, also referred to as sciatic nerve pain, is not an injury or condition itself but rather a collection of well-known symptoms. Several healthcare professionals estimate that up to 80 percent of people may experience some form of back pain throughout their lifetime, including low back pain and sciatica. The purpose of the article below is to discuss the causes and symptoms of low back pain and sciatica as well as demonstrate the treatment approaches to improve health and wellness. �
Contents
Causes and Symptoms of Sciatica
The most common symptoms associated with sciatica include pain and discomfort along the length of the sciatic nerve, tingling sensations or “pins-and-needles” in the feet and toes as well as numbness. The painful symptoms may vary in severity and these may also often become aggravated by sitting for extended periods of time. Sciatica is common in a variety of health issues, however, an estimated 90 percent of cases are due to a herniated disc. Other common causes of sciatica include: �
Lumbar spinal stenosis or the narrowing of the spinal canal in the low back
Spondylolisthesis, a condition where a vertebra slips forward over the vertebra below it
Spinal tumors, which may compress or impinge the sciatic nerve
Infection, which may ultimately affect the spine
Other causes like an injury to the lumbar spine or lower back
Cauda equina syndrome, a rare but serious condition which affects the nerves in the lower part of the spinal cord; it generally requires immediate medical attention, and
Pregnancy, which can affect approximately 50 to 80 percent of pregnant women.
Diagnosis and Treatment of Sciatica
If the sciatica symptoms are moderate and don’t last more than 4 to 8 weeks, it’s referred to as acute sciatica, and immediate medical attention isn’t generally required. As for chronic sciatica which does require immediate medical attention, a complete medical history may help speed up the diagnosis process. Shooting pain down the length of the leg with physical activities generally indicates sciatica.�Imaging tests, such as an X-ray or MRI, may help determine the source of sciatica. �
Acute Sciatica Treatment
Most cases of acute sciatica can be treated utilizing a variety of self-care measures, which include: �
Over-the-counter painkillers, such as ibuprofen,
Stretches and exercises, such as walking
Hot or cold therapy. It is generally helpful to alternate between the two
Not all treatments are suitable for everybody; individuals may need to talk to their healthcare professional. �
Chronic Sciatica Treatment
Treatment for chronic sciatica generally involves a combination of self-care measures and medical treatment, including: �
physical therapy
chiropractic care
cognitive behavioral therapy or CBT
Surgery may be an option if symptoms have not improved with other treatment approaches. Surgical options include: �
Lumbar laminectomy or the widening of the spinal cord in the lower back to reduce pressure on the nerves.
Discectomy, the partial or entire removal of a herniated disc
Depending on the cause of sciatica, a surgeon will go over the risks and benefits of surgery and suggest the best option. �
Stretches and Exercises
As previously mentioned above, stretches and exercises can help improve sciatica symptoms. This allows patients to: �
Alleviate sciatica symptoms on their own
Reduce or prevent the use of drugs and/or medications
Find relief and comfort for sciatica symptoms during flare-ups
Sciatica, or sciatic nerve pain, is a collection of symptoms, rather than a single injury or condition, characterized by pain and discomfort, tingling sensations, and numbness anywhere along the length of the sciatic nerve. The sciatic nerve is the longest and largest nerve in the human body which extends from the lower back, down the buttocks, hips, and thighs, into the legs, knees, and feet. The compression or impingement of the sciatic nerve and irritation due to a herniated disc, among other health issues, can ultimately cause sciatica symptoms and low back pain. – Dr. Alex Jimenez D.C., C.C.S.T. Insight
Low Back Pain
The purpose of the article was to discuss and demonstrate the low back pain associated with sciatica. Sciatica is a collection of symptoms characterized by pain and discomfort, tingling sensation, and numbness. The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues as well as functional medicine articles, topics, and discussions. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 . �
Curated by Dr. Alex Jimenez �
Additional Topic Discussion: Foot Orthotics
Low back pain and sciatica are common health issues which affect many individuals worldwide. However, did you know that chronic pain may be due to foot problems? Health issues originating in the foot may ultimately cause imbalances in the spine, such as poor posture, which can cause the well-known symptoms of low back pain and sciatica. Custom foot orthotics, individually designed with 3-arch support can help promote overall health and wellness by supporting and promoting good posture and correcting foot problems. Custom foot orthotics can ultimately help improve low back pain and sciatica. �
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
�
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download �
* All of the above XYMOGEN policies remain strictly in force. �
If you have never been to see a chiropractor before, you may be wondering what you should expect. You likely have seen a video or heard someone talk about a chiropractic adjustment. Having your back or neck adjusted is definitely a big part of the process�but what else do you need to know? Continue reading to discover the basics of chiropractic and what you have to look forward to.
Contents
�Chiropractic Guide for First Time Patients
Your visit begins with a thorough examination.
Don�t expect to walk through the door of the chiropractor and get your back cracked. Chiropractors are highly-trained professionals that focus on individualized treatments and getting the best possible results. That means getting a clear picture of what your condition is and identifying the ideal treatments before starting with the adjustments.
The chiropractor will conduct a full physical examination to identify your injuries, any movement difficulties, etc. He or she will also ask a number of questions to find out how you feel and why you think you feel the way you do.
You might get an adjustment.
There are a lot of ways to adjust the human body, which are not limited to the standard adjustments you may have seen on TV or in movies. Your chiropractor can adjust not only your back but all of your joints, including knees, ankles, elbows and more. He or she can also use different tools to facilitate adjustments and employ techniques that range from extremely gentle to more forceful.
The adjustment you get will be based on your unique circumstances and the opinion of the chiropractor.
You might not get an adjustment.
It can be surprising to be told by the chiropractor that you won�t get an adjustment during your visit, but it does happen. Your chiropractor may determine that other therapies are better for your current condition. The adjustment may have to wait until your body is ready for it.
You will be offered alternative therapies.
Chiropractors focus on getting the best results for patients, which often means employing supplemental and complementary therapies in addition to chiropractic adjustments. Some common therapies include massage, decompression, electrical stimulation and more. Massage can relax soft tissues to allow for better adjustments, or to help adjustments stay in place. Decompression can increase blood flow to discs. Electrical stimulation can increase healing times.
You always have the final say in your treatment plan.
While the chiropractor will certainly have recommendations on which therapies are best suited to your needs, in the end, it is you who decides what happens. After your examination, the chiropractor will present the recommended options and let you decide if you want to start treatment.
You will experience improvements sooner rather than later.
Many patients are pleasantly surprised by how effective their chiropractic treatments are. Often after the first visit, you will feel less pain, more mobile and more at ease. A good adjustment and supplemental therapies can do a great deal to alleviate discomfort and stiffness. The good news is that each successive visit will only add to the good feelings you experience.
Getting Safe, Highly Effective Treatment
According to the American Chiropractic Association, chiropractic care is one of the safest drug-free, non-invasive treatments available for injuries and degenerative conditions related to your muscles, joints and skeletal system. There are so many benefits, and so few potential downsides, that it is hard to find a reason not to try chiropractic!
Please contact our team today to schedule an appointment with a chiropractor. Discover why so many people choose chiropractic for their healthcare needs.
Chiropractic
Gait Related Lower Back Pain
31 million Americans experience low back pain at any given time. This condition affects many, but finding the exact cause can be a challenge. Chiropractors are spinal specialists that are trained to not only alleviate pain but also find and help correct the cause of the problem.� Prevention is the best cure for any condition/ailment. When someone sees a chiropractor, they not only find relief for their condition, but they learn ways to prevent their symptoms in the future. Using proper exercise and ergonomics, they can ease the discomfort before it starts.
NCBI Resources
A�chiropractor�is the ideal�medical professional�to consult with for any unexplained pain in the musculoskeletal system. They are highly qualified professionals that their specialty is treating conditions like lower back pain and they are very affordable. If you or a loved one have pain in the lower back, gives us a call. We�re here to help!
Most people know that a chiropractor can help with musculoskeletal problems, such as backaches and slipped discs, but many people do not realize that chiropractic can do much more. In fact, chiropractic can even improve your sleep.
Getting enough quality sleep each night is good for your health. Not getting enough sleep can increase your risk of developing obesity, diabetes, cardiovascular disease, and other serious conditions, according to Harvard Medical School.
But what if you allow yourself plenty of time but still have trouble falling asleep or staying asleep?
Chiropractic can help you sleep. In fact, one-third of people who undergo chiropractic adjustment report sleeping better immediately, according to the National Sleep Foundation. The benefits are not just for adults � 40 percent of infants slept better after just one chiropractic session. That is good news, considering one-fourth of babies experience sleep problems and when babies don�t sleep, nobody else in the house sleeps either!
Contents
3 Ways Chiropractic Can Improve Your Sleep
1. Pain relief
Pain can prevent you from getting a decent night�s sleep, especially if you have chronic pain that lasts for three or more months. In fact, one study shows that 20 percent of people with chronic pain experience at least one symptom of insomnia, compared with only 7.4 percent of people without chronic pain. Chiropractic can alleviate pain associated with backaches, headaches, whiplash, migraines and more to help you get the sleep you need.
Chiropractic can also help you sleep by providing a drug-free approach to pain relief. Many prescription pain relievers, such as those containing morphine, oxycodone, and opioids, can make you drowsy throughout the day. Daytime drowsiness might cause you to take naps throughout the day, which can interfere with your ability to sleep at night. The drug-free approach of chiropractic allows you to stay alert all day and sleep well at night.
Extensive research presented by the National Institutes of Health shows that chiropractic is as effective as other treatments, such as drugs, at relieving pain. Perhaps the best part of chiropractic is that the research shows that pain relief lasts up to 18 months of treatment � which means you can remain pain-free without using drugs for a year and a half.
2. Relieves muscle tension and stress
Stress can prevent you from falling asleep or sleeping well. Stress can also cause uncomfortable muscle tension that interferes with sleep. The National Sleep Foundation lists muscle tension as one of the three signs that you are under too much stress to sleep. Tension and poor sleep can be a vicious cycle, as tension can cause poor sleep and poor sleep can cause tension.
Chiropractic often includes massage to melt away painful muscle spasms. These therapeutic massages are relaxing, which helps you unwind enough to fall asleep. Chiropractic also stimulates blood flow, which helps bring oxygen to your brain and other parts of your body.
Your chiropractor may also be able to suggest gentle exercises, stretches and relaxation techniques to help you relax tense muscles and quiet your stressed-out mind.
3. Suggested sleeping positions
Your chiropractor may offer suggestions on sleeping positions and other ideas about how you can sleep better. Your chiropractor may discourage you from sleeping on your stomach if you are having back pain, for example, or be able to recommend a particular pillow or mattress to help you sleep better at night.
For more information on how chiropractic can improve your sleep, talk with your chiropractor. You may start sleeping better as soon as your first visit.
Reduce *IMBALANCE & PAIN* with Functional Orthotics | El Paso, Tx
Foot Orthotics
Foot Dysfunction�can very easily cause a domino effect that extends all the way to the back. The feet are the foundation of the body and when there is a problem with the way they function it can cause the entire body to shift out of alignment.
Recipe Ingredient Substitutions
Here is a guide to give your kitchen a healthy makeover! Here are tips to help you create a healthy environment for your home, and these substitutions will help you make delicious allergy-friendly�dishes.
NCBI Resources
We live in an extremely sleep-deprived society. According to the�Centers for Disease Control�(CDC), 1 in 3 people in the United States does not get enough sleep on a regular basis. Additionally, the�National Sleep Foundation�reports that 45 percent of adults in the US admit that insufficient or poor sleep has had an impact on their daily activities at least once in the previous week. What�s more, about a third of the people who said they slept the number of hours that doctors recommend reported experiencing poor sleep quality.
Functional hallux limitus is a differential diagnosis from structural hallux limitus. Functional hallux limitus, or FnHL, is identified by the movement deficiency of the first metatarsophalangeal joint, found between the bones of the foot and of the toes, during gait. The first metatarsophalangeal, or MTP, joint reveals regular movements throughout an open kinetic chain evaluation. Radiographs may demonstrate a tiny amount of dorsal spurring of the joint, however, the joint doesn’t always necessarily demonstrate pervasive degenerative joint disease as an ultimate source of the health issue. �
Structural hallux limitus, or SHL, is identified by structural adaptations of the first MTP joint which restrict ordinary movement from happening in the first place. These changes can be acute, with very limited dorsiflexion mobility, )hallux rigidus) or minor, (hallux limitus) leading to small changes in dorsiflexion of the first MTP joint. When normal foot insertion during propulsion is interrupted during MTP joint movement to prevent functioning of the foot construction through maximal hallux dorsiflexion over the effect of the windlass mechanism, the small changes can become considerably significant. In SHL, movement is going to be disrupted during open-and-closed-kinetic string actions. Pain and joint mobility is ultimately connected with any movement of the first metatarsophalangeal, or MTP, joint of the foot and of the toes. �
�
Although the health issue itself is generally considered to be asymptomatic, the signs and symptoms of this condition can also be associated with a variety of other common pathologies of the foot, including heel pain, lesser metatarsal pain, Morton’s neuroma, Achilles tendon pain, and retrocalcaneal enthesitis as well as posterior tibial tendon dysfunction and postural alignment abnormalities of the lower back, or lumbar spine, and the lower extremities, including sciatica. Research studies have demonstrated that the prevalence of the disorder is much higher among the symptomatic population. �
Contents
Identifying Functional Hallux Limitus
Functional hallux limitus is unfortunately still considered to be a rare health issue which often goes unaddressed. Outcomes for many different foot disorders will be jeopardized if treatment doesn’t address functional hallux limitus. Foot and postural abnormalities have a basis in poor function of stabilization and the windlass mechanism of the foot structure through the plantar fascia. By way of instance, if this stabilizing effect is not present, it’s hypothesized that a range of foot disorders may result as normal foot function which cannot be achieved without appropriate purpose of the plantar fascia. Addressing the proper function of the foot has a high level of predictability for healing mechanically induced foot pain and this remains the goal of footbed and shoe modifications as the primary focus of orthotic intervention throughout many years in medicine. �
Functional hallux limitus, or FnHL, is commonly diagnosed by a healthcare professional by detecting gait patterns and utilizing pedobarographic evaluations, which often require a qualified and experienced doctor or pricey equipment. The time consuming evaluations are also seldom able to be performed in the clinical setting. These challenges may probably contribute to the underdiagnosis of FnHL, or functional hallux limitus. Moreover, many healthcare professionals have also identified an FnHL evaluation demonstrated by the stiffening of large toe movement when implementing a loading force from the stage of maximal displacement to the first ray, especially when holding the foot and the toes in a neutral position. �
When hallux dorsiflexion at the MTP joint is ultimately evaluated, there is a stiffening or bending movement of the MTP joint, when roughly equal power is applied to the hallux as is applied to the first metatarsal head. Healthcare professionals have demonstrated how several evaluations associated with visual gait evaluations have demonstrated that 72 percent of patients with a pronated midtarsal joint also had a positive evaluation for functional hallux limitus. Approximately 66 percent of patients with normal midtarsal joint motion had a negative test result. Healthcare professionals believe this to be a reliable test for the diagnosis of abnormal foot function. These type of evaluation have been conducted on asymptomatic individuals. �
Functional Hallux Limitus and Sciatica
Functional hallux limitus is often commonly asymptomatic, therefore, it is rarely recognized as the functional inability of the first metatarsophalangeal, or MTP, joint to dorsiflex during gait. Normal movement is present in this joint during non-weight-bearing evaluations. Because the joint controls the pivot from which the entire body moves forward during each step, this disturbance in function, when repeated multiple times on a regular basis, can ultimately affect foot and postural biomechanics. It can also cause and aggravate many underlying health issues, including low back pain and sciatica. When functional hallux limitus is addressed in an orthotic treatment plan, 77 percent of long-term postural problem patients demonstrate 50 percent to 100 percent improvement in their overall health and wellness, among other health issues. �
As soon as the diagnosis for FnHL has been made, the healthcare professional is going to want to address this health issue with the best treatment approach. Because this is a fundamental derangement of movement of the first MTP joint, conservative treatment is often successful and can be easily demonstrated in the foot evaluation. Surgery is seldom indicated. This is in contrast to structural hallux limitus, or SHL, which often does require surgical interventions to restore pain-free assortment of motion. FnHL is generally a misalignment of the first MTP joint in which the first metatarsal is dorsally displaced, restricting the average movement of the proximal phalanx in the first metatarsal head. It’s essential for a healthcare professional to accurately diagnose functional hallux limitus to follow-up with the proper treatment. �
Because of the multifaceted nature of functional hallux limitus, understanding the health issue is ultimately fundamental for healthcare professionals and patients alike. Fortunately, there are many treatment approaches for functional hallux limitus depending on the nature of the pathology. As described above, functional hallux limitus is demonstrated as a normal range of hallux dorsiflexion during non-weight-bearing, however, there is a considerable decrease in hallux dorsiflexion. Functional hallux limitus has also been demonstrated to cause sciatica. – Dr. Alex Jimenez D.C., C.C.S.T. Insight
Fibromyalgia Magazine
The purpose of the article was to discuss the differential diagnosis in sciatica with a variety of health issues, including functional hallux limitus as well as structural hallux limitus, among others. Accurate diagnosis and proper treatment are important for the treatment of functional hallux limitus and low back pain or sciatica. The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues as well as functional medicine articles, topics, and discussions. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 . �
Curated by Dr. Alex Jimenez �
Additional Topic Discussion: Severe Sciatica
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. Your spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of sciatica, or sciatic nerve pain. Sports injuries or automobile accident injuries are often the most frequent cause of painful symptoms, however, sometimes the simplest of movements can have these results. Fortunately, alternative treatment options, such as chiropractic care, can help ease sciatic nerve pain, or sciatica, through the utilization of spinal adjustments and manual manipulations, ultimately improving pain relief. �
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
�
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download �
* All of the above XYMOGEN policies remain strictly in force. �
Back pain and everything that comes with it can really take you out of it. It, being life, that you don’t get to participate in, because you have to anticipate how, whatever the activity may be, is going to affect your back. So you just skip activities altogether. No more! You have options and chiropractic care is one of them. Because back pain can occur due to a variety of health issues, diagnosis is essential to create the correct treatment plan.
Doctor of chiropractic Alexander Jimenez specializes in treating back pain. A few patients share their back pain experiences and how it affected their lives. They describe how Dr. Jimenez has helped them improve their overall health and wellness with chiropractic care. Patients highly recommend Dr. Jimenez and his staff as the back pain specialist.
The feet are the foundation of the body; they support its weight. If there is not enough support, various parts of the body, especially the spine become exposed to added stress, this can lead to:
Back, hip, knee and foot pain
Poor posture and abnormal foot movement
Sports performance compromised
Greater risk of injury
An abnormal walk leads to imbalances throughout the knees, ankles, and feet, and can eventually cause pain throughout the body, and yes you guessed it the back.
Foot Levelers Kiosk
The Kiosk helps guide patients in selecting the best custom-made orthotics for their condition and lifestyle. It’s�
Fast:�Push the Start button and the scanner begins.
Easy to use:�User-friendly easy touch screen.
Engaging:�Videos explain the importance of healthy feet and the benefits of custom-made orthotics.
Cloud-based:�Results can be securely accessed from anywhere.
Comprehensive:�Easily retrieve previous scans to compare them to new scans and see the difference.
The Foot Levelers Kiosk helps you. It saves time so you can spend more time living your life.
Intake Form
This is the intake form that is used to set up a custom orthotics treatment plan.
Low Back Pain
When there are problems with the feet, it can cause problems through the legs and all the way to the spine. This can cause the ankle to pronate, meaning it rolls inward. This alters the way the�bones of the foot�line up which extends through the tibia, or shin bone.
The spine is the foundation for the body, supporting the spinal cord and the limbs. When there is a problem with any of the limbs it can affect the spine and vice versa. The intricate network of tendons, ligaments, and nerves work together within the incredible machine that is the human body.
When you visit a doctor office for a medical concern, the healthcare professional will perform a variety of diagnostic procedures to determine the source of your symptoms. The doctor will review the patient’s current symptoms and medical history as well as the patient’s results from a physical evaluation to diagnose an injury and/or an underlying condition. �
Once the healthcare professional has reviewed the patient’s current symptoms, medical history, and results from the physical evaluation, the doctor will make a list of the probable injuries and/or underlying conditions which may be causing the patient’s symptoms. This is the differential diagnosis. A differential diagnosis refers to the process of differentiating between two or more health issues which share similar signs and symptoms. The doctor will perform additional tests or assessments to rule out specific injuries and/or underlying conditions in order to reach a final diagnosis and follow-up with treatment. �
The differential diagnosis generally varies depending on the health issue. A variety of health issues can cause common symptoms which may make it challenging to determine the source of the pain and discomfort. Sciatica is a collection of symptoms rather than a single condition which can be a frequent symptom for many health issues. Sciatica is characterized as pain, tingling sensations, and numbness, due to compression or impingement along the length of the sciatic nerve. �
Sciatica commonly occurs when an underlying health issue results in the compression or impingement of the sciatic nerve in the lower back. A bulging or herniated disk is one of the most common causes of sciatica. However, a variety of other underlying health issues can cause sciatica and determining what causes sciatica is necessary for proper treatment. In part 2 of this article, we will demonstrate the differences between sciatica and other health issues with similar painful symptoms.� �
Contents
Sacroiliac Joint Dysfunction
The sacroiliac joint is generally attributed to causing between 15 to 30 percent of chronic low back pain cases. Irritation, swelling, or inflammation of one or more sacroiliac joints is commonly referred to as sacroiliac joint dysfunction, sacroiliac joint disease, or sacroiliitis. Moreover, sacroiliac joint dysfunction or disease may cause sacroiliitis. �
Sacroiliac joint dysfunction is characterized as a sharp, stabbing pain which radiates from the pelvis and hips, down into the lower back or lumbar spine and throughout the legs. Patients might experience tingling sensations or numbness. Every person experiences symptoms of SI joint dysfunction differently and the signs can vary from person to person, depending on the source of the sacroiliac joint dysfunction. Common signs and symptoms of SI joint dysfunction include: � �
low back pain
pain in the buttocks, hips, and pelvis
pain in the groin
painful symptoms in the SI joints
pain when standing from a sitting position
stiffness
burning sensations
weakness
numbness
pain radiating down into the thighs and legs
feeling like the legs may buckle and not support the weight of the body
Because the joints are situated deep within the human body, it often makes it difficult for healthcare professionals to diagnose the health issue. The healthcare professional may perform a variety of tests to diagnose SI joint dysfunction, such as provocative tests, injections, and/or imaging tests like X-rays, MRIs, and CT scans to be able to follow-up with treatment. �
Gluteus Tendinopathy
Gluteus medius tendinopathy (GMT), also known as dead butt syndrome (DBS), is a painful health issue caused by the inflammation of the tendons in the gluteus medius muscle. Although GMT most commonly affects athletes, it can also affect people starting a new exercise program, during an intense workout routine or due to an underlying health issue. �
The common symptoms associated with gluteus medius tendinopathy, or GMT, include pain and discomfort, stiffness, and weakness in the hip or buttocks region. The painful symptoms can generally worsen throughout weight-bearing exercises, such as walking, running, and/or climbing. For many people, the pain and discomfort associated with dead butt syndrome, or DBS, can radiate down the hips, buttocks, and legs or thighs, similar to sciatica and hamstring tendinopathy. Sciatica is a collection of symptoms characterized by pain and discomfort, tingling sensations, and numbness along the sciatic nerve. �
Many patients diagnosed with GMT also report experiencing painful symptoms when lying in bed on the affected hip or buttocks region as well as pain, discomfort, stiffness, and weakness throughout the night and upon rising in the morning. Moreover, if DBS progresses past the initial stage, the hip bursa may become inflamed, another health issue known as trochanteric bursitis, which may also cause swelling, tenderness, redness or warmth in the hip, among other health issues. �
When the tendon becomes inflamed due to an injury or an aggravated underlying condition, such as fibromyalgia, the gluteus medius can fail to trigger appropriately, hence why the “dead butt” term is utilized as an alternate name for this health issue. People sitting down for extended periods of time may cause their hip flexors to become tight and failure to stretch before physical activities can cause DBS. If left untreated, gluteus medius tendinopathy (GMT), or dead butt syndrome (DBS) may lead to complications, including trochanteric bursitis, IT band syndrome, and plantar fasciitis, among other health issues.
Ischiogluteal Bursitis
A bursa is a fluid-filled sac which functions by helping to decrease friction around the joints. However, when the fluid inside a bursa becomes infected or irritated due to excessive movement, the bursa can become swollen, causing pain and discomfort, ultimately making it difficult to move the affected joint. This health issue is commonly known as bursitis. �
Although bursitis generally occurs in the knee, called �housemaid�s knee�, or in the elbow, called �tennis elbow�, it can occur in any joint in the human body. When it occurs in the buttocks, it is known as ischiogluteal bursitis. In this instance, the bursa is found between the ischial tuberosity at the base of the pelvis and the tendon of the hamstring muscle. �
The symptoms of ischiogluteal bursitis can be similar to those of hamstring tendonitis. Several of the most common symptoms associated with ischiogluteal bursitis include pain and discomfort as well as tenderness in the buttocks region. Painful symptoms may frequently occur when stretching the hamstring muscle or flexing the knee against resistance. �
The symptoms of ischiogluteal bursitis can also be similar to those of sciatic nerve pain. Sciatica is a collection of symptoms characterized by pain, discomfort, tingling sensations, and numbness. While the symptoms of ischiogluteal bursitis may be similar to those of sciatica, sciatic nerve pain occurs when the sciatic nerve is irritated, compressed or impinged due to an injury and/or an aggravated condition. Ischiogluteal bursitis caused by an infection can ultimately be followed by a fever.
The three most common causes of bursitis are generally due to injury or trauma, infection, and crystal deposits. Trauma can include repetitive movements or a blow to the region. By way of instance, the injury could occur due to a fall. Occasionally, trauma or injury can cause blood to leak into the bursa, causing ischiogluteal bursitis, or any other type of bursitis. �
Infection septic bursitis, or an infection in the bursa, generally occurs in the joints which are located closer to the surface of the human body. This health issue frequently affects men, however, it does not seem to be a leading cause of ischiogluteal bursitis. Crystal deposits can also form around the joints when there is too much uric acid in the human body. People with gout can develop bursitis because it is also caused by crystal deposits and it shares many similar symptoms with bursitis. �
Proximal Hamstring Injury
Hamstring injuries are several of the most common types of injuries among athletes. These makeup for the most days or even weeks missed each year amongst AFL football players. The majority of partial or complete tears include either the hamstring muscle belly or the distal musculotendinous junction. However, a proximal hamstring injury is ultimately uncommon. In the total hamstring injury spectrum, it makes up for under 10 percent of hamstring injuries, among other health issues. �
The proximal hamstring tendon can become injured through progressive stretching or through sudden and intense contraction when the hip is forcefully flexed over an extended knee. In younger patients with an average proximal hamstring tendon, this can occur through sprinting or hurdling, however, the most common athletes affected in this instance involves waterskiiers who fall forward with an extended knee. In elderly patients, proximal hamstring injuries occur through a different type of trauma, such as slipping on a wet surface or even doing the “splits” inadvertently. �
Proximal hamstring injuries could include complete tendon ruptures or incomplete/partial tears. In young patients, the bone together with the tendon is frequently avulsed or fractured in the pelvis or the ischium. In older patients, the tendon usually avulses or tears from the bone of the ischium at its attachment point. Occasionally, the tendon may tear in its midsubstance, leaving a stump of tendon still attached to the bone. Frequently this type of injury is referred to as a partial tear. �
Adductor Injuries
An adductor muscle strain is characterized as an acute injury to the muscles of the groin. Although a variety of muscles in the human body can be injured, the adductor longus, medius, magnus, and the gracilis are some of the most well-known muscle groups to be affected by an injury.� According to healthcare professionals, atrains are muscle tears which generally occur due to the powerful contraction of the muscles against resistance, often during what is referred to as an eccentric load. �
An adductor muscle strain occurs during acute muscle contraction, such as during kicking, pivoting or skating. Factors which can predispose a person to injury include failure to properly stretch or warm up and fatigue from overuse. The risk of experiencing an adductor strain increases with sports involving strides like sprinting, soccer, and hockey. Sports with repeated movements like football, martial arts, and gymnastics can also substantially increase the risk of experiencing an adductor strain. Struggling to warm up, stretch or be properly conditioned can also cause an adductor muscle strain. �
The symptoms associated with an adductor muscle strain most commonly involve a variety of painful symptoms which are most frequently related to other types of muscle strain. Common symptoms of an adductor muscle strain include a sudden onset of pain and discomfort, occasionally accompanied by the feeling of a pop in the inner section of the thigh as well as the inability to maintain action after the first onset of painful symptoms. Because an adductor strain may cause groin or hip pain, many patients and healthcare professionals may confuse this type of injury with sciatica or sciatic nerve pain. �
Adductor tendinopathy is generally characterized as pain on palpation of the adductor tendons, adduction of the legs and/or of the injured leg. Pain can develop gradually or cause a sudden and severe, sharp pain. A swelling or a lump may also be experienced from the adductor muscle(s), stiffness at the groin area or an inability to contract or extend the adductors. In acute cases, exercises and physical activities will be restricted because the tendon can’t sustain repeated tensile loading. �
The adductors are triggered in many sports such as, running, soccer, horse riding, gymnastics, and swimming. The repetitive movements and the continuous change of direction in physical activities activate the adductor tendon, which makes athletes more prevalent to develop adductor tendinopathy and other groin injuries. Other causes of adductor tendinopathy can include over-stretching of the adductor tendons or a sudden increase in training, especially in the type of training. �
Adductor tendinopathy may occur due to considerable leg length discrepancy which affects gait pattern. Poor or abnormal movement patterns may also overly stress the adductor tendons. Muscular length gaps, strength imbalances or muscular weakness in the thoracic or the abdominals can also be powerful in developing adductor tendinopathy. Other causes include a failure to properly warm-up, inactivity, fatigue, obesity, age-related weaknesses, genetics or degeneration. �
Spinal Tumors
Most back pain is caused by other health issues. However, when painful symptoms are caused by spinal tumors, it’s essential for a healthcare professional to perform an accurate diagnosis to follow-up with proper treatment. There are three types of tumors which can cause back pain; vertebral column tumors, intradural-extramedullary tumors, and intramedullary tumors. �
Tumors in the spine can cause back pain when the growth weakens the bone, which can ultimately result in spinal fractures, compression or impingement of the nerves, and spinal distress. Spinal tumor symptoms may ultimately be similar to those of other well-known health issues, including sciatica, especially if the spinal tumor irritates the sciatic nerve.� Furthermore, the following symptoms below may ultimately be commonly associated with a spinal tumor, including: �
Neck pain or back pain followed by neurological health issues, such as tingling sensations, weakness, and numbness in the upper and lower extremities as well as abnormal bladder and bowel habits.
Painful symptoms, particularly in the morning
Severe pain with manipulation of the affected region of the spine
Painful symptoms which do not decrease with rest
Back pain and other associated symptoms, such as loss of appetite, nausea, vomiting, weight loss, fever, or chills.
If any tumor is located in the spine and if there’s no other known cancer, a healthcare professional may perform a series of comprehensive evaluations and additional testing of all common organs in which cancer can develop. Because many vertebral column tumors originate from cancer in another organ, the ultimate goal of spinal tumor treatment is to: � �
Regulate severe painful symptoms which occur with these spinal tumors by removing pressure on the nerve roots
Maintain neurological structure and function by removing pressure on the spinal cord
Correct structural and functional instability in the spine by correcting the unstable spine with a spinal fusion
�
As previously mentioned, when you seek medical attention for a health issue, it’s essential for the healthcare professional to accurately diagnose your medical concern in order to follow-up with the proper treatment. A differential diagnosis is a list of possible injuries and/or underlying conditions which may be causing symptoms. Sciatica, a group of signs and symptoms commonly characterized by pain, discomfort, tingling sensations, and numbness, is a well-known health issue which can have many differential diagnosis. – Dr. Alex Jimenez D.C., C.C.S.T. Insight
Fibromyalgia Magazine
The purpose of the article was to discuss the differential diagnosis in sciatica with a variety of other health issues. Accurate diagnosis and proper treatment are important for sciatic nerve pain management. The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues as well as functional medicine articles, topics, and discussions. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 . �
Curated by Dr. Alex Jimenez �
Additional Topic Discussion: Severe Sciatica
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. Your spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of sciatica, or sciatic nerve pain. Sports injuries or automobile accident injuries are often the most frequent cause of painful symptoms, however, sometimes the simplest of movements can have these results. Fortunately, alternative treatment options, such as chiropractic care, can help ease sciatic nerve pain, or sciatica, through the utilization of spinal adjustments and manual manipulations, ultimately improving pain relief. �
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
�
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download �
* All of the above XYMOGEN policies remain strictly in force. �
Most people think that chiropractic care is something to do if they experience back or neck pain, and rightly so, as these are the most common reasons people visit a chiropractor. But chiropractic does way more than that.
Chiropractic helps with a variety of ailments from headaches to digestive issues and addresses problems like sports injuries, work injuries, arthritis, and injury prevention. Regular trips to the chiropractor are a good idea. Here are three important benefits that you can get from regular chiropractic adjustments.
Contents
What Taking an Active Role in Your Health Can Do
Chiropractic is a holistic kind of care that has a chiropractor serve as a guide as your own body uses its own innate ability to heal itself. When you see a chiropractor on a regular basis, even if there is not a pressing problem, you are taking an active role in assuring you are in the best health you can be.
Chiropractic adjustments correct misalignments in the spine that may be blocking nerve function and causing pain to muscles, joints, and ligaments. When you are adjusted by a chiropractor, these blocks are removed and you are able to move and function better than you did before.
A chiropractor can also provide you with nutritional and lifestyle advice, including exercises you can do between visits that will improve strength, flexibility, and various other functions. Without your active participation and cooperation, the success of chiropractic diminishes, so you can feel good about taking responsibility for your own health.
Improved Neurological Conditions
The spine connects to several different areas of the body, including the brain. Adjustments have been shown to increase blood flow to the brain, which in turn improves function in many areas.
The brain and spine are frequent communicators, and the nervous system is often the “wires” that are used to assure proper communication. Stress can trigger misalignments and the breakdown in these systems. Chiropractic is able to identify when stress is placed on these nerves and has even been shown to increase white blood cell count in some patients.
Eliminated Need for Pain Medications
One of the cornerstones of chiropractic is that it does not rely on pain medications or surgery in order to treat pain. Instead, it is designed to trigger your body’s own ability to heal itself without drugs and surgery.
Through methods such as manual adjustment, massage, and electrical muscle stimulation, chiropractic strengthens communication throughout the body in order to help it heal naturally. Chiropractors use important diagnostic tools such as x-rays, laboratory analysis, and reviewing your health history to pursue a treatment plan.
Conversely, when misalignments are allowed to build up over time, there is a greater risk that you will be tempted to seek out pain medications in order to mask the pain temporarily. While occasional use of these is okay, over time they can build up in the system and damage liver function, so it is important to find other methods of pain management whenever possible.
In closing, chiropractic benefits the overall health of people that utilize it consistently. So if you’re ready to experience these benefits yourself, just pick up the phone and give us a call. We’re here to help!
Benefits of Custom-Made *ORTHOTICS* | El Paso, Tx
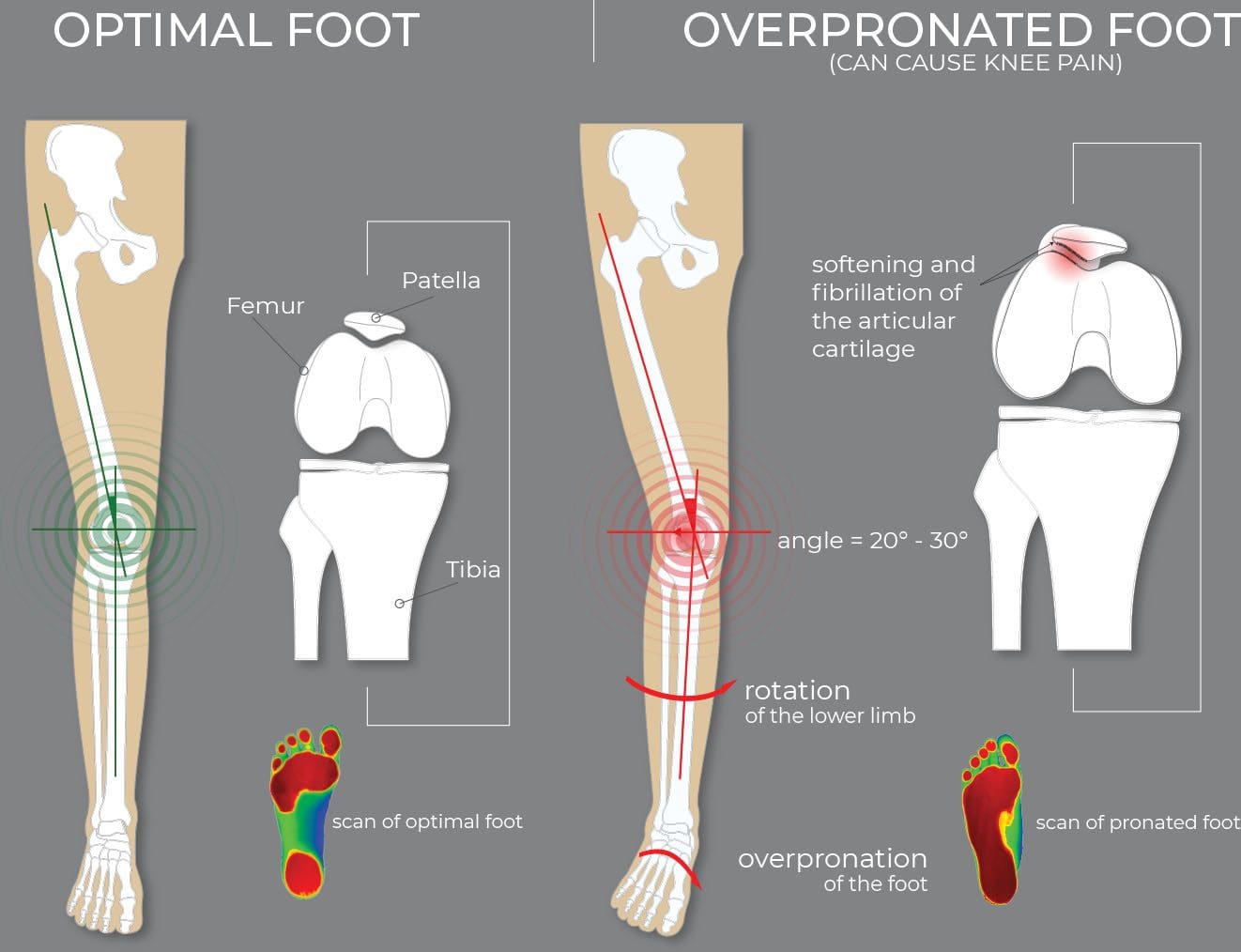
Overpronation
99% of the population has some degree of foot pronation.Foot pronation occurs when the foot/feet roll inwards.
People with excessive pronation may experience inward rotation of the knee and forward tilting of the pelvis, which causes pain in the:
Feet
Knees
Hips
Back
Shoulders
Neck
Foot Orthotics
Foot Dysfunction�can very easily cause a domino effect that extends all the way to the back. The feet are the foundation of the body and when there is a problem with the way they function it can cause the entire body to shift out of alignment.
Pronation & Misalignment
For the foot to function correctly, there must be a significant degree of pronation. However, excessive pronation, or overpronation, can cause injury and damage to the foot and ankle. It creates the arch in the foot to flatten, and the ligaments, tendons, and muscles under the foot overstretch.
NCBI Resources
Chiropractic seeks to find the cause of the conditions it is used to treat, including pain, instead of just treating symptoms. Because of this, the chiropractor will work to find the cause of the pain.
When you visit a doctor office for a medical concern, the healthcare professional will perform a variety of diagnostic procedures to determine the source of your symptoms. The doctor will review the patient’s current symptoms and medical history as well as the patient’s results from a physical evaluation to diagnose an injury and/or underlying condition. �
Once the healthcare professional has reviewed the patient’s current symptoms, medical history, and results from the physical evaluation, the doctor will make a list of the probable injuries and/or underlying conditions which may be causing the patient’s symptoms. This is the differential diagnosis. A differential diagnosis refers to the process of differentiating between two or more health issues which share similar signs and symptoms. The doctor will perform additional tests or assessments to rule out specific injuries and/or underlying conditions in order to reach a final diagnosis and follow-up with treatment. �
The differential diagnosis generally varies depending on the health issue. A variety of health issues can cause common symptoms which may make it challenging to determine the source of the pain and discomfort. Sciatica is a collection of symptoms rather than a single condition which can be a frequent symptom for many health issues. Sciatica is characterized as pain, tingling sensations, and numbness, due to compression or impingement along the length of the sciatic nerve. �
Contents
Sciatica
The characteristic symptoms of sciatica can range from mild to severe where the sciatic nerve pain is described as a dull ache to pain or as a sharp, searing, or electric-like pain. Symptoms differ from one person to another. By way of instance, some people may experience shooting pain while some people may only experience tingling sensations and/or numbness. � The common symptoms of sciatica, also known as sciatic nerve pain, include: �
Radiating pain along the length of the sciatic nerve
Discomfort, tingling sensations, and numbness
Muscle weakness in the lower back, hips, legs, and feet
A combination of pain, discomfort, tingling sensations and numbness
Painful sensations on one or both sides of the human body
Sciatica commonly occurs when an underlying health issue results in the compression or impingement of the sciatic nerve in the lower back. A bulging or herniated disk is one of the most common causes of sciatica. However, a variety of other underlying health issues can cause sciatica and determining what causes sciatica is necessary for proper treatment. In part 1 of this article, we will demonstrate the differences between sciatica and other health issues with similar painful symptoms. �
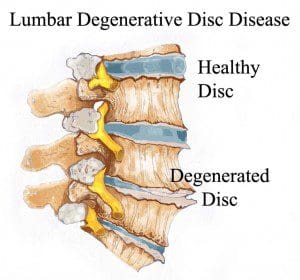
Degenerative Disc Disease
The intervertebral discs also referred to as spinal discs or intervertebral fibrocartilage, provide the padding required between the vertebrae of the spine. The intervertebral discs are an elastic structure made from fibrocartilage tissue. The intervertebral discs cushion the stress of the spine, bears weight, and also helps the spine bend and flex. �
As people age, repeated daily stresses on the backbone and occasional trauma and/or injuries, including minor, undetected health issues, may ultimately damage the intervertebral discs in the back. Changes caused by damage may include: �
Decreased fluid: The intervertebral disks of a healthy young adult are made up of around 90 percent fluid. With age, the fluid material decreases, causing the disc to become thinner. The distance between each vertebra, in turn, becomes smaller and it makes them even less effective to function as a cushion or shock-absorber.
Disc structure: Small tears or cracks can become larger in the outer layer of the disc. The soft and gelatinous material from the inner part may push through the disc, causing a bulging or ruptured disc. The disc may break into fragments.
If the vertebrae have less padding between them, the backbone also becomes less stable. To compensate, the human body builds osteophytes, or bone spurs, small bony structures which develop along the edge of bones. These structures can compress or impinge the spinal cord or nerve roots. These changes can cause painful symptoms and lead to fatigue. �
The pain and discomfort can range from mild to severe and it may often be debilitating. It may result in osteoarthritis along with pain and stiffness. When degenerative disc disease affects the low back or the lumbar spine, the pain may radiate down the buttocks, hips, and thighs, into the knees and feet. There might also be tingling sensations and numbness, the collection of symptoms known as sciatica, caused due to the compression or impingement of the sciatic nerve. �
Radiculopathy
The spine consists of 33 bones, best known as vertebrae, which shield the spinal cord from experiencing injury or trauma. Each vertebra is cushioned from each other with an intervertebral disk. This shields the vertebrae from rubbing over each other. When trauma or injury does occur, these intervertebral discs can become damaged and cause the compression or impingement of a nerve. Depending on which nerve is compressed, an individual can experience a variety of painful symptoms, including sciatica or sciatic nerve pain. Individuals can also develop radiculopathy seemingly without a cause. � Radiculopathy can ultimately develop due to a variety of injuries and/or aggravated conditions, including: �
Herniated discs, or when a disc protrudes, irritating the nerve root
Sciatica
Degenerative disc disease
Bone spurs
Tumors in the spine
Osteoarthritis or spinal arthritis
Spinal stenosis, or the narrowing of the spinal canal
Compression fractures
Spondylolisthesis, or when a vertebra slides over the vertebra beneath it
Scoliosis brought on by an abnormal curve in the backbone
Diabetes, caused by altered nerve blood flow
Cauda equine syndrome, or a condition where nerve root compression affects the pelvic organs and lower extremities
� Additional risk factors for developing radiculopathy include: � �
Aging
Being overweight or obese
Poor posture
Improper lifting methods and techniques
Repetitive movements
A family history of degenerative bone health issues
Individuals may experience painful symptoms in the low back, hips, and legs when nerve compression or impingement occurs in the lumbar spine or low back region of the spine. Lumbar radiculopathy is also commonly referred to as sciatica or sciatic nerve pain. Sciatica symptoms include pain, tingling sensations, and numbness along the length of the sciatic nerve. Occasionally, the nerve roots in charge of controlling the bowel and bladder may become irritated, resulting in bowel or bladder incontinence as well as loss of control. Other generalized symptoms associated with lumbar radiculopathy include: �
Sharp pain extending from the low back to the foot
Sharp pain with sitting or coughing
Weakness or numbness in the lower extremities
Tingling sensations and numbness in the lower extremities
Hypersensitivity, or sensitivity and reflex alterations
Sharp pain radiating into the upper extremities
Intense pain and discomfort with movements of the head and/or neck
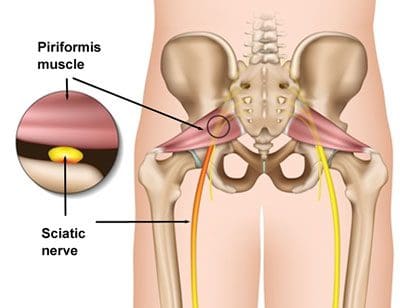
Piriformis Syndrome
People with sciatic nerve pain, or sciatica, will often report experiencing pain and discomfort, tingling sensations, and numbness anywhere along the length of the sciatic nerve. The sciatic nerve is the longest and largest nerve in the human body, which travels from the lower back into the hip and buttocks, down into the thighs, knees, legs, and feet. People with sciatica, or sciatic nerve pain, may also experience another common health issue known as piriformis syndrome. � �
Piriformis syndrome can cause sciatica when a piriformis muscle spasm irritates the sciatic nerve. The piriformis is a muscle which extends from the front of the sacrum, the triangle-shaped bone between the two hipbones in the pelvis, across the sciatic nerve and into the top of the femur, the large bone in the upper leg. When an injury or underlying condition causes the piriformis muscle to compress or impinge the sciatic nerve due to inflammation, the end result is piriformis syndrome. �
Sciatica is the most common symptom associated with piriformis syndrome. Other common symptoms associated with piriformis syndrome include tingling sensations and numbness, muscle tenderness, pain and discomfort while sitting or engaging in physical activities, and difficulty sitting comfortably. The pain and discomfort will generally manifest on one or both sides of the hips, buttocks, and thighs, where it will then radiate down the back of the leg, knees, and feet. �
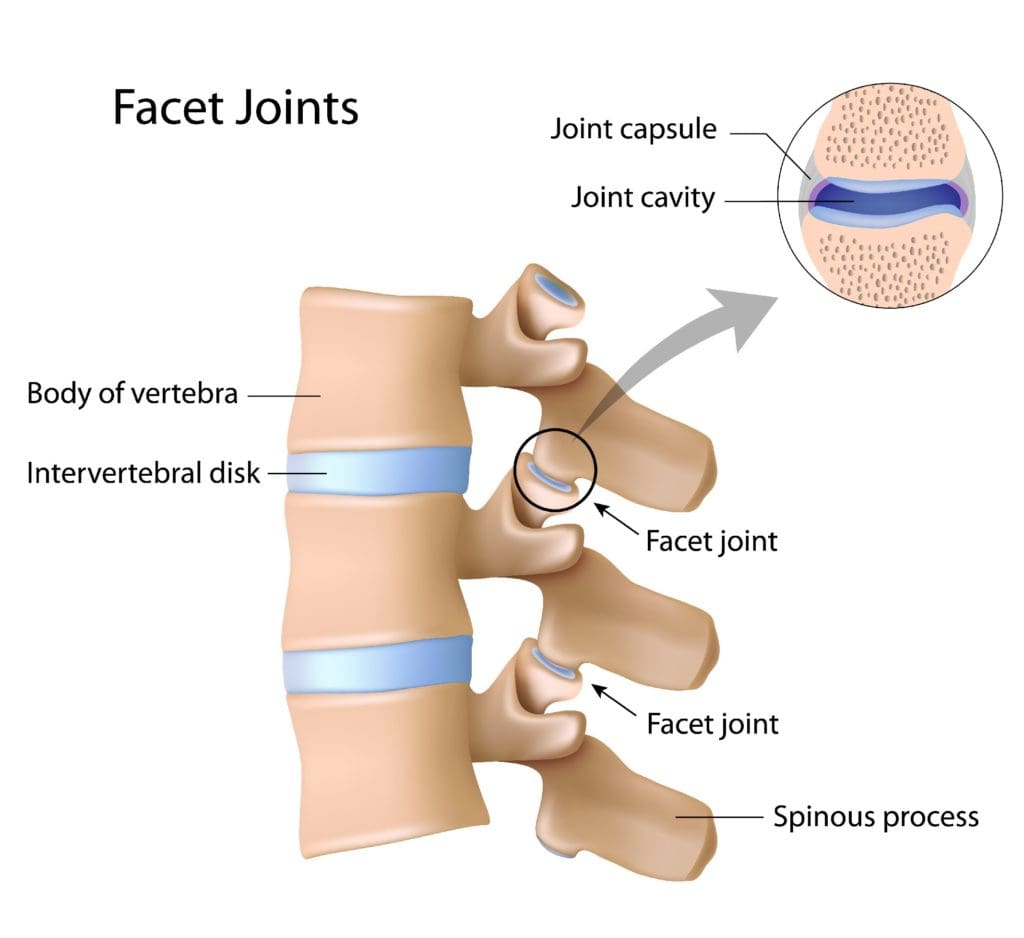
Facet Arthropathy
The facet joints are the joints which are found behind the spine to counterbalance the intervertebral discs found between the vertebrae of the spine. Over time, the natural aging process can cause the facet joints to deteriorate or wear down, causing facet joint arthritis or facet arthropathy. Individuals with facet arthropathy will experience low back pain as a dull ache on one or both sides of the lower back or lumbar spine which can worsen with standing, twisting, or bending backward. �
However, unlike the well-known signs and symptoms of sciatica, caused by the compression or impingement of the sciatic nerve in the lower back, facet arthropathy signs and symptoms generally don’t radiate down the buttocks, hips, and thighs, or into the legs and feet. However, the facet joint, in the same way as any other joint which has arthritis, can also become enlarged and add pressure on nerve roots, causing pain and discomfort to radiate down into the lower extremities. �
Facet arthropathy may cause bone spurs, tiny bone outgrowths. Bone spurs can decrease the distance available between nerve roots, causing a health issue known as spinal stenosis. Spinal stenosis may cause pain, weakness, and numbness on the buttocks, hips, and thighs. It’s frequently associated with other health issues which could lead to facet arthropathy. The natural aging process is frequently considered to be one of the most common indirect sources of facet arthropathy. Other health issues which can ultimately affect the facet joints and cause facet arthropathy include: � �
Osteoarthritis: Degeneration of joint cartilage and underlying bone, generally during middle age
Facet joint degeneration: Wear and tear on the facet joint brought on over time due to aging
Facet joint injury: Trauma to the facet joints caused by an impact, such as a fall or automobile accident
Synovial cyst: A fluid-filled sac which develops in the spine, generally as a result of aging
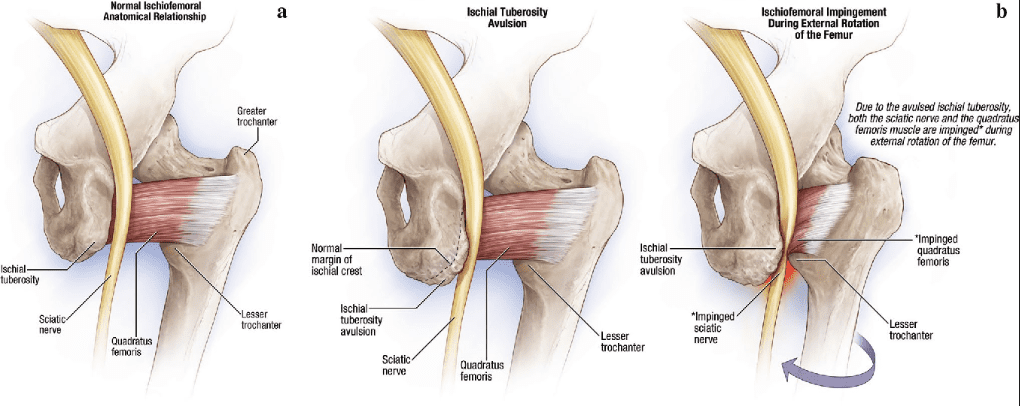
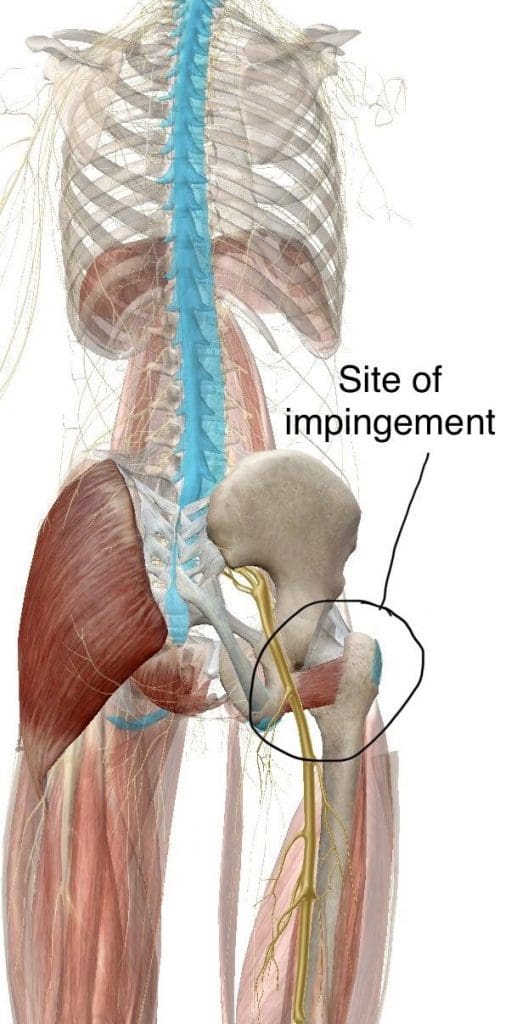
Ischiofemoral Impingement
Ischiofemoral impingement syndrome is a condition which occurs when the ischium and the head of the femur experience excessive amounts of pressure, generally due to trauma, overuse, or surgery. The trauma, overuse, or surgery, can also cause bone spurs to grow along the ischium. This gives the bones an abnormal shape which makes them no longer fit together. �
Because these don�t fit properly together anymore, the bones may then start to rub against each other during movement causing friction which can damage the joint and cause painful symptoms. As friction worsens, the pain may also increase and restrict movements. The symptoms for ischiofemoral impingement may vary and treatment can also depend on symptoms. �
�
The main symptom of ischiofemoral impingement syndrome is hip pain. The pain from this underlying health issue may start off as mild, however, it can often worsen as the rubbing of the bones causes more damage and inflammation. Ischiofemoral impingement syndrome can feel similar to a pulled hamstring but the pain is generally closer to the buttocks like sciatica. �
Ischiofemoral impingement occurs due to excess friction between the top of the thighbone, or the femur, and the hip bone, or the ischium. These two bones generally touch in a way which allows them to move without causing pain, however, trauma, overuse, and surgery can ultimately damage the bone. Moreover, if the damage causes a bone spur to grow on the hip or femur, as described above, the extra piece of bone�can cause more damage and make the condition worse.
When you seek medical attention for a medical concern, it’s fundamental for the healthcare professional to accurately diagnose your health issue in order to follow-up with the proper treatment. A differential diagnosis is a list of possible injuries and/or underlying conditions which may be causing symptoms. Sciatica, a collection of symptoms commonly characterized by pain, discomfort, tingling sensations, and numbness, is a well-known health issue which can have many differential diagnosis. – Dr. Alex Jimenez D.C., C.C.S.T. Insight
Fibromyalgia Magazine
The purpose of the article was to discuss the differential diagnosis in sciatica with a variety of other health issues. Accurate diagnosis and proper treatment are important for sciatic nerve pain management. The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues as well as functional medicine articles, topics, and discussions. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 . �
Curated by Dr. Alex Jimenez �
Additional Topic Discussion: Severe Sciatica
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. Your spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of sciatica, or sciatic nerve pain. Sports injuries or automobile accident injuries are often the most frequent cause of painful symptoms, however, sometimes the simplest of movements can have these results. Fortunately, alternative treatment options, such as chiropractic care, can help ease sciatic nerve pain, or sciatica, through the utilization of spinal adjustments and manual manipulations, ultimately improving pain relief. �
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
�
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download �
* All of the above XYMOGEN policies remain strictly in force. �
IFM's Find A Practitioner tool is the largest referral network in Functional Medicine, created to help patients locate Functional Medicine practitioners anywhere in the world. IFM Certified Practitioners are listed first in the search results, given their extensive education in Functional Medicine




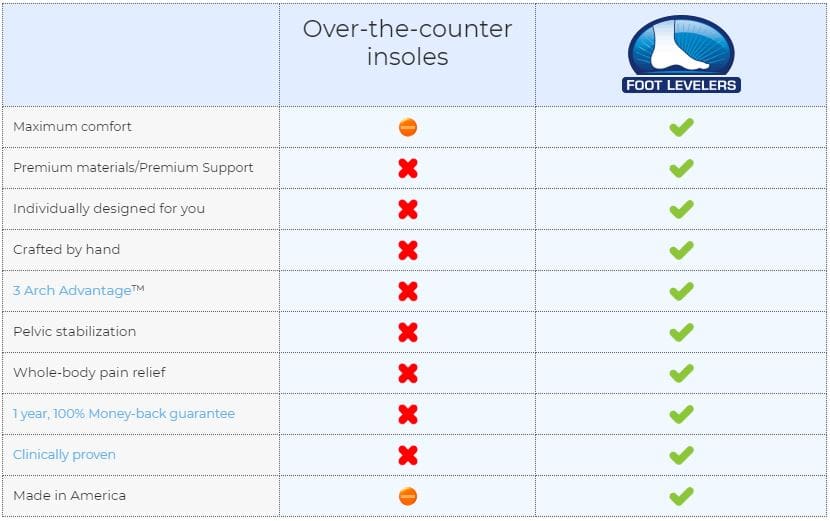 Store-bought orthotic insoles are cheap but the cost to you and your health can be very high. Understanding the differences between over the counter and custom made orthotics is important. Foot Levelers customized orthotics provide the highest-quality and most effective orthotic out there.
Foot Levelers custom orthotics are individually designed for your feet. This achieves a balanced foundation and a stabilized pelvis. These orthotics are handmade based on 3D scans or casts, and your doctor�s examination.
Over-the-counter insoles do cost less, but they only support one arch in the foot. When only one arch is supported, the structure can collapse, and that’s when problems can begin in other parts of the body. Over-support of one arch can cause pain and symptoms, instead of relieving them.
Store-bought orthotic insoles are cheap but the cost to you and your health can be very high. Understanding the differences between over the counter and custom made orthotics is important. Foot Levelers customized orthotics provide the highest-quality and most effective orthotic out there.
Foot Levelers custom orthotics are individually designed for your feet. This achieves a balanced foundation and a stabilized pelvis. These orthotics are handmade based on 3D scans or casts, and your doctor�s examination.
Over-the-counter insoles do cost less, but they only support one arch in the foot. When only one arch is supported, the structure can collapse, and that’s when problems can begin in other parts of the body. Over-support of one arch can cause pain and symptoms, instead of relieving them.






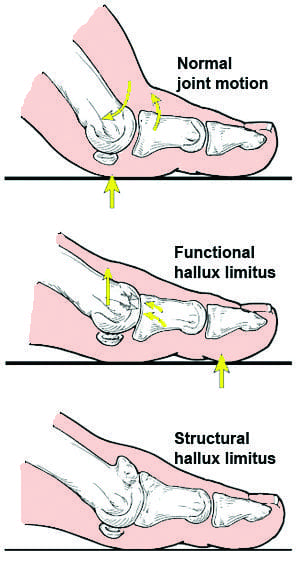 �
�

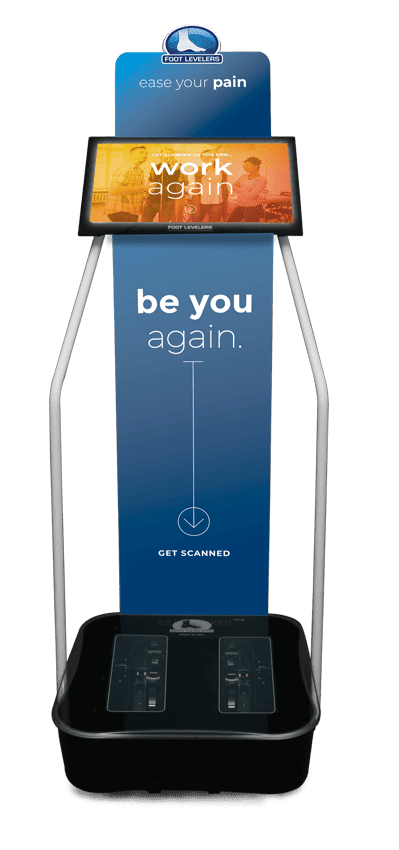

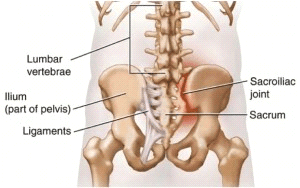
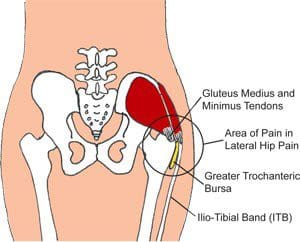
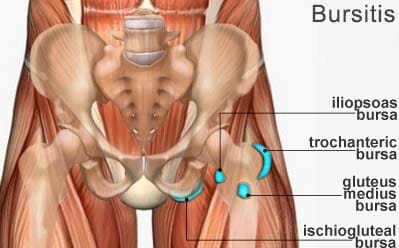
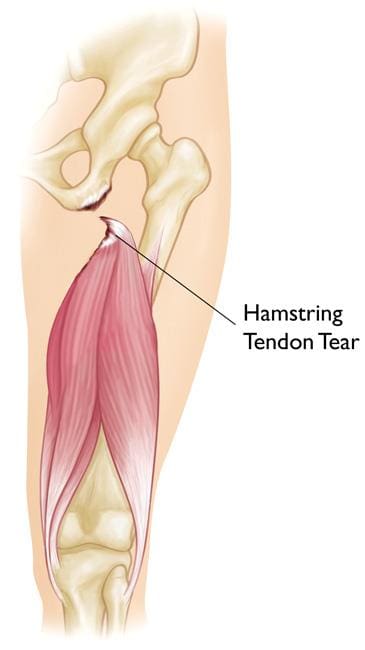
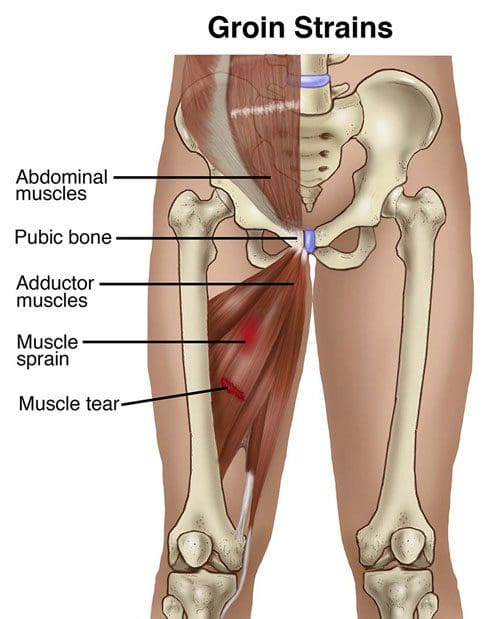
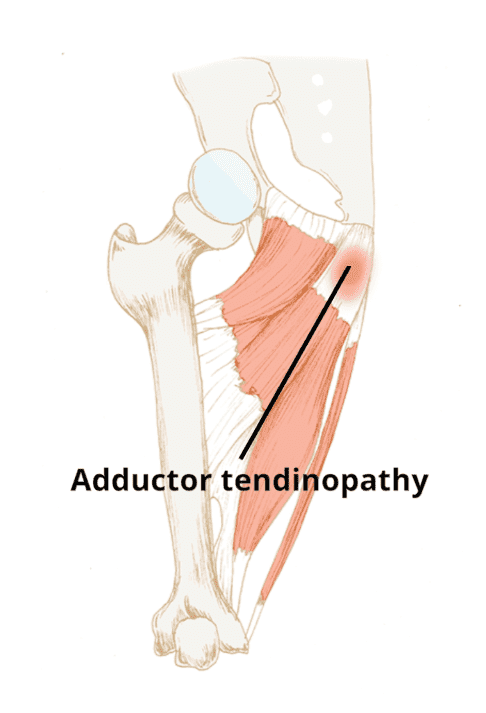
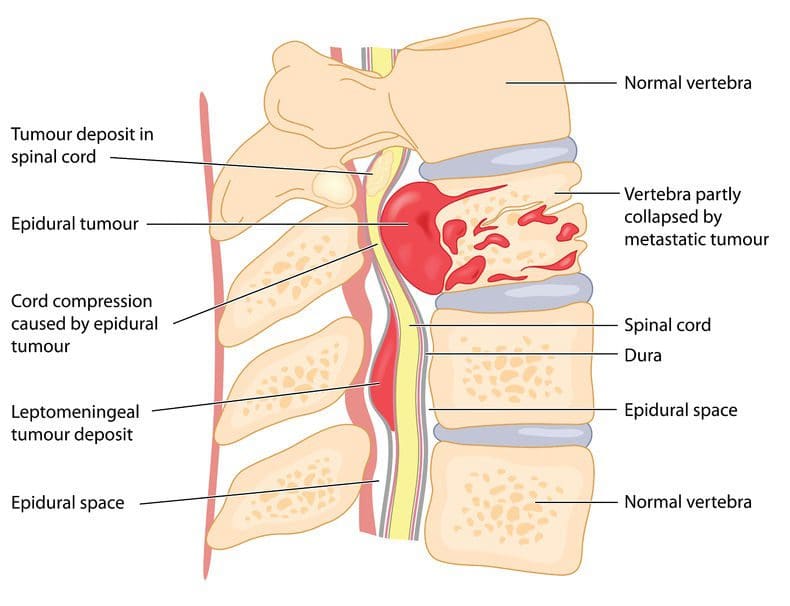


 Foot pronation occurs when the foot/feet roll inwards.
People with excessive pronation may experience inward rotation of the knee and forward tilting of the pelvis, which causes pain in the:
Foot pronation occurs when the foot/feet roll inwards.
People with excessive pronation may experience inward rotation of the knee and forward tilting of the pelvis, which causes pain in the: