The calves are extremely important to the lower portions of the legs as they allow the individual to move around and help stabilize the entire body. The calves have two muscles: the gastrocnemius and the soleus, which provide plantarflexion and stability at the ankle joint. These two muscles work together to ensure that the upper body’s weight doesn’t lean forward too much to let the individual fall. However, when many people overuse the calf muscles constantly, it can cause the muscle fibers in the lower legs to be overstretched and develop small nodules known as trigger points to cause referred pain to the calves and the ankles that can affect a person’s ability to walk, run, or jump. Today’s article looks at the soleus muscle, how trigger points affect the muscle, and different methods to reduce pain while managing trigger points along the calves. We refer patients to certified providers that incorporate various techniques in the lower body extremities, like lower leg and calve pain therapies correlating to trigger points, to aid many people dealing with pain symptoms along the soleus muscles, causing muscle cramps and issues along the ankles. We encourage and appreciate each patient by referring them to associated medical providers based on their diagnosis when it is appropriate. We understand that education is an excellent way when asking our providers intricated questions at the patient’s request and understanding. Dr. Alex Jimenez, D.C., only utilizes this information as an educational service. Disclaimer
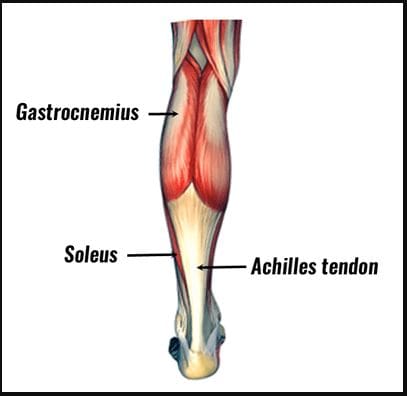
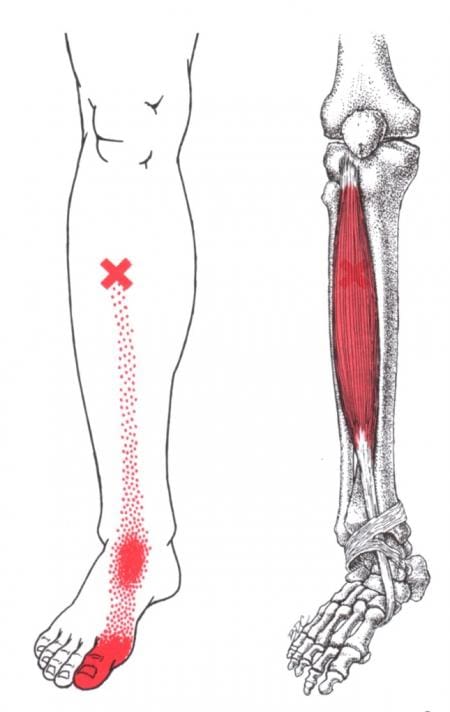
What Is The Soleus Muscle?
Do you feel a cramping sensation in your calves? Do your feet hurt when you are walking, jumping, or running? Or are you experiencing tightness in your calve muscles? When a person is experiencing calf pain in their legs, it could be trigger points causing overlapping issues in the soleus muscles. As stated earlier, the calves have two muscles: the gastrocnemius and the soleus. The gastrocnemius makes up the round shape of the calves and is a complex, superficial muscle. At the same time, the soleus is a large, flat muscle deep within the gastrocnemius and forms the calcaneal tendon (Achilles tendon), which can be mistaken as a nerve for the ankle. Studies reveal that the soleus muscle works together with the gastrocnemius, and these muscles constitute a plantar flexor. The soleus is part of a group of muscles known as antigravity muscles. It acts like a skeletal muscle and helps maintain good posture in the body to prevent the sheer body weight from falling forwards at the ankle when standing.
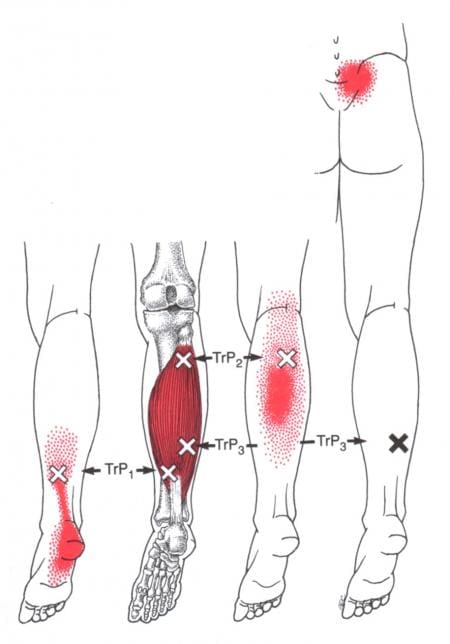
How Do Trigger Points Affect The Soleus Muscle?
The calves are essential for the movement and mobility functions of the body. When the calve muscles have been overused to repetitive movements or have been dealing with trauma, it can cause the muscle fibers to tear and develop trigger points along the calves causing referred pain to travel down the ankles. Trigger points along the soleus muscle are tricky to diagnose since they cause referred pain to the surrounding muscles. “Myofascial Pain and Dysfunction,” written by Dr. Janet G. Travell, M.D., mention that, unlike the latent trigger points that can affect the gastrocnemius, active trigger points can affect the soleus muscle and cause tenderness referred to the heel. When muscle tenderness affects the heels, it can cause trigger points to mimic chronic conditions like plantar fasciitis. Additional studies reveal that musculoskeletal impairments like trigger points in the soleus muscle could potentially lead to biomechanical changes in the ankle. When trigger points cause hypersensitivity spots in the skeletal muscle taut band, it can restrict the soleus muscle to allow ankle dorsiflexion. Individuals with active soleus trigger points are prone to develop low back pain due to ankle dorsiflexion restriction, leading to them leaning over and lifting objects improperly.
Trigger Point Of The Week: Soleus – Video
Have you been dealing with pain traveling down from your calves to your ankles? Experiencing stiffness around your ankles? Or do you feel like you are about to fall? These pain-like issues are associated with trigger points along the soleus muscles along the calves. The soleus is one of the two muscles (the other is the gastrocnemius) that make up the calves and acts as a skeletal muscle to help maintain good posture in the body. When trigger points affect the soleus muscle, they can cause referred pain to the calves and mimic chronic issues like plantar fasciitis at the ankles. The video above explains where the soleus muscles are located, deep within the gastrocnemius, where the trigger points are causing referred pain to the ankles, and where they are in the soleus muscle fibers in the calves. Even though trigger points can cause referred pain in the affected muscle fibers of the body, they are treatable and can be managed through different methods to help the calves.
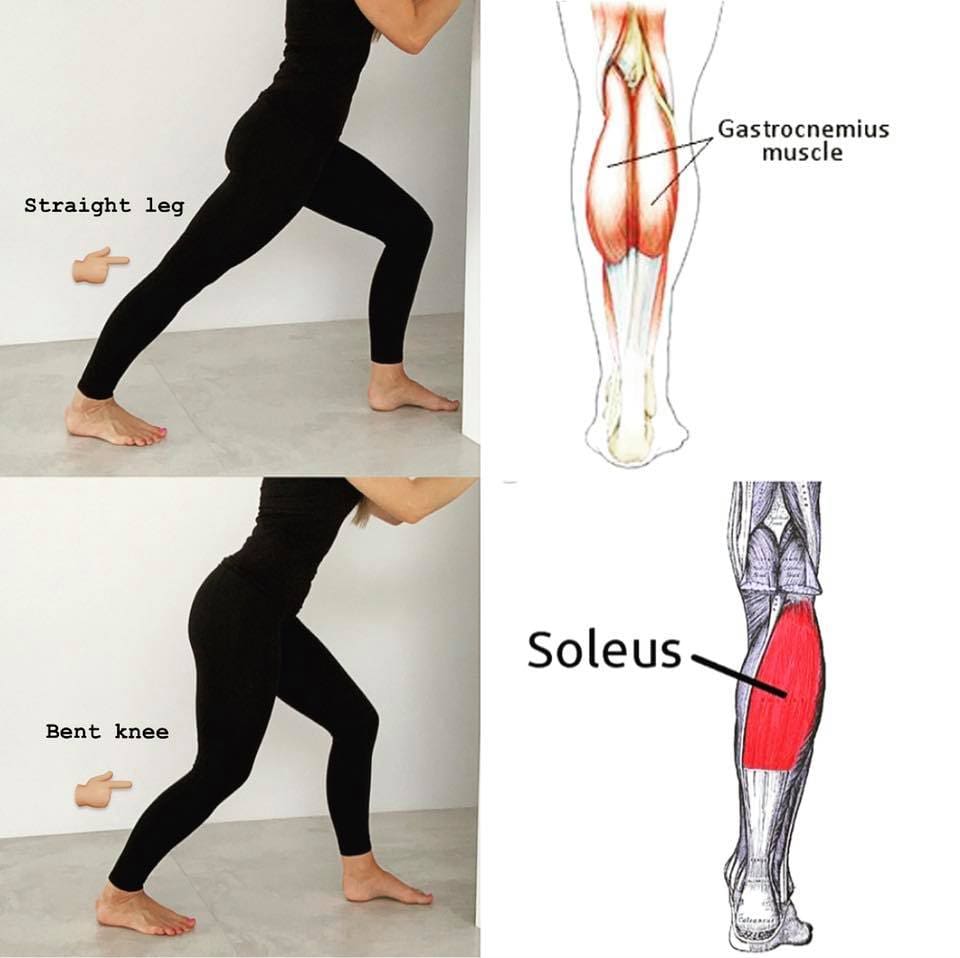
Different Methods To Reduce Pain And Manage Trigger Points In The Calves
Even though trigger points can affect the soleus muscles and cause referred pain to the ankles, different methods can reduce the pain and help manage trigger points in the calves. Studies reveal that when there are multitudes of therapies that can help manage trigger points, it can help reduce the pain affecting the soleus muscle by optimizing muscle function and preventing the development of chronic pain syndromes. Besides therapies that target trigger points, corrective actions and stretches can help release the pain from the soleus muscle. Incorporating calf stretches, having a correct posture when sitting down, wearing long loose socks while sleeping, and wearing low heels can prevent the soleus muscle from shortening and help manage trigger points from re-occurring in the calves. When people start to take care of their calve muscles, it can help their mobility and stability at the ankles.
Conclusion
As part of the calf muscles and an antigravity muscle, the soleus works with the gastrocnemius to help with stability and plantarflexion to the ankles. This muscle is deep within the gastrocnemius and helps maintain good posture in the body that prevents a person from falling forward at the ankles when standing. When the soleus muscles have been overused, they can develop tiny knots in the muscle fibers known as trigger points that can cause referred pain to the ankles. Trigger points along the soleus muscles can cause overlapping features in the calves by restricting ankle dorsiflexion, causing individuals to develop low back pain associated with leaning forward. However, various therapies can manage trigger points through multiple treatments that can reduce the pain and allow stability back to the calves, enabling the individual to continue walking without feeling pain.
References
Binstead, Justin T, et al. “Anatomy, Bony Pelvis and Lower Limb, Calf – Statpearls – NCBI Bookshelf.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 29 May 2022, https://www.ncbi.nlm.nih.gov/books/NBK459362/.
Travell, J. G., et al. Myofascial Pain and Dysfunction: The Trigger Point Manual: Vol. 2:the Lower Extremities. Williams & Wilkins, 1999.
Valera-Calero, Juan Antonio, et al. “Prediction Model of Soleus Muscle Depth Based on Anthropometric Features: Potential Applications for Dry Needling.” Diagnostics (Basel, Switzerland), U.S. National Library of Medicine, 7 May 2020, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7277950/.
Various muscles help support the lower legs in the body and allow movement to the host. The lower extremities include the hips, pelvis, thighs, legs, knees, and feet. At the same time, the various muscles help provide support and stability to the upper body’s weight and incorporate mobility and movement for the entire body to go from one location to another. The legs have two sections connected with the knees; the upper portion has the hips and thigh muscles, while the lower legs have the calve muscles, shin muscles, and the Achilles tendon. The calve muscles have two groups of muscles, and when the calve intense exercises, or normal factors have overused muscles may potentially lead to muscle cramps associated with trigger points. Today’s article examines one of the calve muscles known as the gastrocnemius, how the calves are affected by trigger points and corrective actions to prevent muscle cramps in the calves. We refer patients to certified providers that incorporate various techniques in the lower body extremities, like lower leg and calve pain therapies correlating to trigger points, to aid many people dealing with pain symptoms along the gastrocnemius muscles, causing muscle cramps. We encourage and appreciate each patient by referring them to associated medical providers based on their diagnosis when it is appropriate. We understand that education is an excellent way when asking our providers intricated questions at the patient’s request and understanding. Dr. Alex Jimenez, D.C., only utilizes this information as an educational service. Disclaimer
What Are The Gastrocnemius Muscles?
Have you been dealing with walking from one place to another? Do your calves feel hard or tense with the slightest touch or movement? Or are you feeling excruciating pain in your calves that make it hard to move? These pain-like symptoms are indicators of trigger points associated with the calves affecting the gastrocnemius muscles. The calves are mostly referred to as the posterior portion of the lower legs responsible for the foot and ankle plantarflexion while also engaging in activities like running or jumping. The two muscles that make up the calves are the gastrocnemius and the soleus. The gastrocnemius is a complex, superficial muscle fundamental to good posture or walking. This muscle has a casual relationship with the lower body as it affects hip movement and the lumbar area of the spine. The gastrocnemius provides a round shape for the calves to form and narrows down to the ankles, where it forms a tendon.
How Are The Calves Affected By Trigger Point Pain?
Since the gastrocnemius provides the round shape to form the calves when the muscles have become overused or injured in a sports activity, it can cause the individual to have limited mobility. Studies reveal that a tear in the gastrocnemius muscles can implicate lower leg trauma and affect the muscle function to plantar flex the foot at the ankle joint and reduce flexion on the leg to the leg knee joint. When it comes to the development of trigger points along the gastrocnemius muscles that are affecting the calves, according to “Myofascial Pain and Dysfunction,” written by Dr. Janet Travell, M.D., the book states that latent trigger points along the gastrocnemius may cause individuals to complain about calf cramps on the legs, however, when the trigger points are active, the individual is aware of the calf pain and would complain about experiencing pain in the back of their knees. The book also mentioned that trigger points associated with the gastrocnemius muscle could be mistaken for other conditions like plantar foot pain or radiculopathy in the sacroiliac joints. When trigger points affect the calf muscles, it can lead to mobility issues and causes people to be unstable when walking.
Trigger Point Of The Week: Gastrocnemius Muscle- Video
Have you been dealing with calf pain when walking for a short distance? Do your calf muscles ache or tense up when you put slight pressure when stepping down? Or do you feel that your calf muscles are hard when resting? Many of these issues affecting the calves are associated with trigger points affecting the gastrocnemius muscles. The gastrocnemius muscle is one of the muscles that make up the calves in the lower legs. This complex, superficial muscle provides a round shape to the calves and can become overused in various activities that can cause a person to have limited mobility. When the gastrocnemius muscle is overused, it can develop tiny nodules in the muscle fibers known as trigger points that mimic other conditions affecting leg mobility. The video above shows where this complex muscle is in the calves and where the trigger points are located in the muscle fibers. Trigger points along the affected muscle can cause referred pain while mimicking other conditions that can often confuse people about what they are feeling. All is not lost, however, as trigger points are treatable and can be managed through various treatments.
Corrective Actions To Prevent Muscle Cramps On The Calves
When the calf muscles like the gastrocnemius are causing symptoms of pain and muscle cramps due to trigger points, there are various treatments and corrective actions that can prevent muscle cramps from causing more issues in the legs and even help manage trigger points from re-forming in the muscle fibers. Some corrective actions that can help improve calve function are gently flexing the foot at the ankle joint to allow the calf muscles to tract and retract to reduce swelling and pain. Other correction actions that people should incorporate to prevent muscle cramping in the calves when they are sitting is to gently rock in a chair to reduce prolonged immobility to the calves and increase blood flow. Studies reveal that dry needling and other various treatments can help reduce muscle stiffness in the gastrocnemius and improve muscle strength in the calves.
Conclusion
The calves are part of the legs that allow plantarflexion at the ankle joint. Known as the gastrocnemius muscle, it makes up the calves’ round shape. The gastrocnemius muscle is complex and superficial as it forms a rounded shape at the calves and narrows down at the ankles. However, when the muscle has been through various activities and is constantly overstretched, it can affect a person’s walking mobility and develop tiny nodules known as trigger points. Trigger points in the gastrocnemius muscle can cause referred pain in the calve muscles and mimic conditions like plantar foot pain to the feet. Fortunately, various treatments and corrective actions can prevent referred pain from re-occurring in the calves and bring mobility back to the legs so a person can continue to their destination.
References
Albin, S R, et al. “The Effect of Dry Needling on Gastrocnemius Muscle Stiffness and Strength in Participants with Latent Trigger Points.” Journal of Electromyography and Kinesiology : Official Journal of the International Society of Electrophysiological Kinesiology, U.S. National Library of Medicine, 9 Oct. 2020, https://pubmed.ncbi.nlm.nih.gov/33075711/.
Binstead, Justin T, et al. “Anatomy, Bony Pelvis and Lower Limb, Calf.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 29 May 2022, https://www.ncbi.nlm.nih.gov/books/NBK459362/.
Bordoni, Bruno, and Matthew Varacallo. “Anatomy, Bony Pelvis and Lower Limb, Gastrocnemius Muscle.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 26 Apr. 2022, https://www.ncbi.nlm.nih.gov/books/NBK532946/.
Nsitem, Virginia. “Diagnosis and Rehabilitation of Gastrocnemius Muscle Tear: A Case Report.” The Journal of the Canadian Chiropractic Association, U.S. National Library of Medicine, Dec. 2013, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3845475/.
Travell, J. G., et al. Myofascial Pain and Dysfunction: The Trigger Point Manual: Vol. 2:the Lower Extremities. Williams & Wilkins, 1999.
The ankles and the lower legs have a casual relationship by allowing movement to the foot that causes an up-and-down motion. The lower leg has various muscles and tendons that surround the shin bone and allows the feet to take the body from one location to another. The peroneal muscles in the legs allow ankle stability to ensure that the weight from the host’s body doesn’t cause overload to the legs and ankles. However, factors like obesity, trauma, or overexerting can cause the peroneal muscles to be inflamed and develop issues like weak ankles or trigger points that can cause referred pain to the ankles and affect how a person walks. Today’s article examines the peroneal muscles, how weak ankles correlate with trigger points, and ways to strengthen the ankles while managing trigger points. We refer patients to certified providers that incorporate various techniques in the lower body extremities, like lower leg and ankle pain therapies correlating to trigger points, to aid many people dealing with pain symptoms along the peroneal muscles, causing weak ankles. We encourage and appreciate each patient by referring them to associated medical providers based on their diagnosis when it is appropriate. We understand that education is an excellent way when asking our providers intricated questions at the patient’s request and understanding. Dr. Alex Jimenez, D.C., only utilizes this information as an educational service. Disclaimer
The Peroneal Muscles On The Ankles
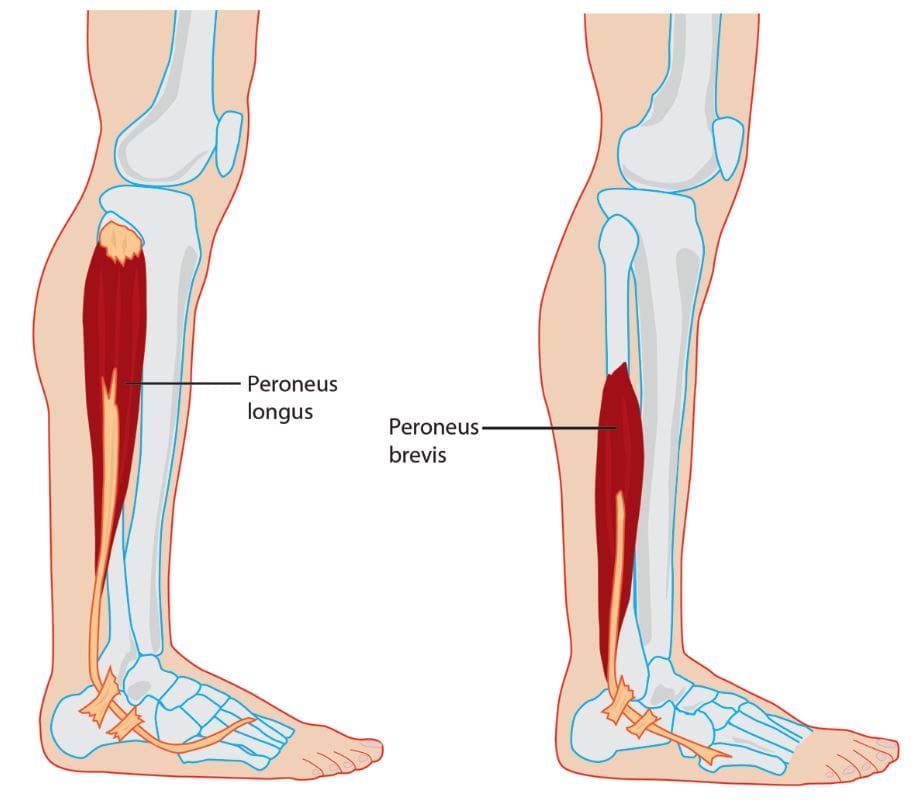
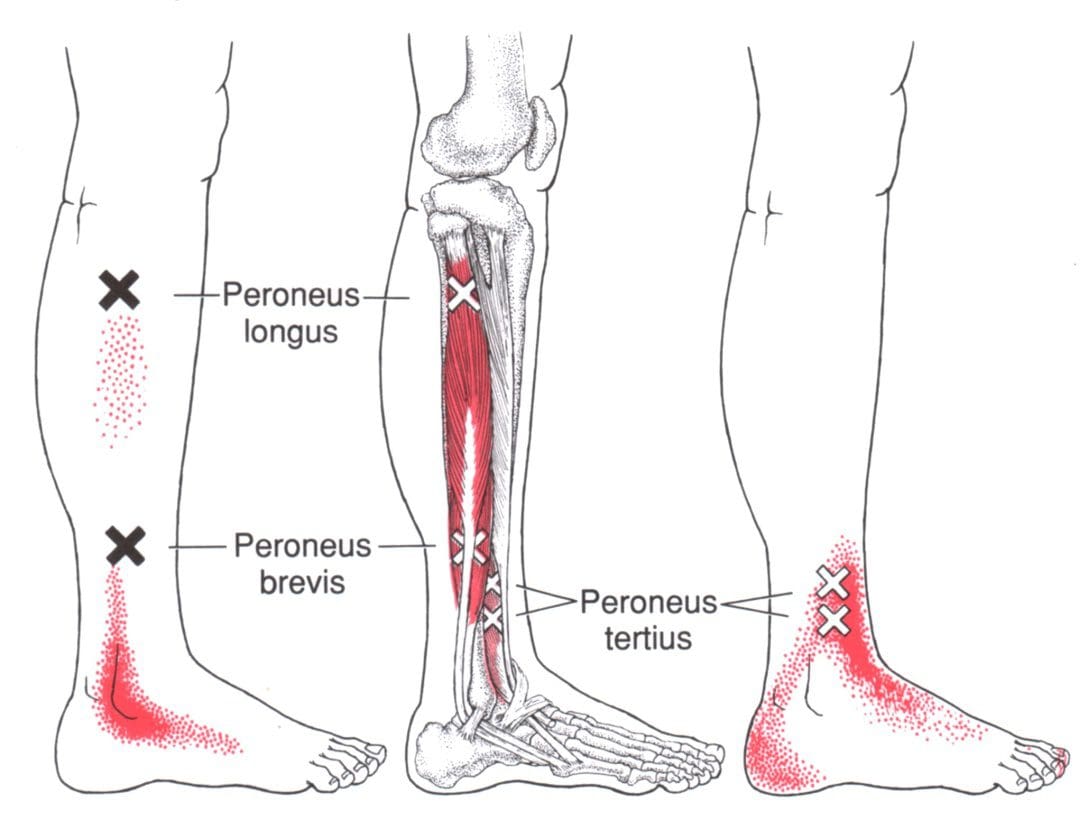
Have you been experiencing pain when walking around constantly? What about feeling a sharp or dull ache in the back or side of your legs? Or do you feel like falling when you are just standing around? Many people experiencing these issues on their legs and ankles could be dealing with trigger points along the peroneal muscles in the ankles. The peroneal muscles consist of two muscles in the lateral compartment of the lower legs: the peroneus longus and peroneus brevis. The peroneus longus is an important long muscle in the lower legs as it is at the top of the fibula and then runs down the outer leg while connecting to the foot. One of the primary functions of the peroneus longus is allowing plantarflex and evert the foot at the ankle. This means that the peroneus longus helps provide motor strength and range of motion to the ankles.
The peroneus brevis is one of the shorter peroneal muscles in the legs that go down to the ankles and provides assistance to allow eversion to the foot and plantarflexion to the ankles. This shorter muscle is important since the ankle joint is relatively mobile and needs stability from the surrounding ligaments and muscles. These two muscles work together for ankle stability when walking and positioning when the body is moving. Studies reveal that depending on a person’s environment, the peroneal muscles allow support and stability to the ankle in various positions. A good example is if the foot is placed in a sloped position, the peroneal muscles and the surrounding ligaments help stabilize the ankle so it won’t induce pain, causing the individual not to fall over.
Weak Ankles & Trigger Points
When factors like obesity, trauma, or injuries begin to affect the lower half of the body, it can cause instability in the legs and cause the surrounding muscles, tendons, and ligaments to be overstretched, take on more of an overload to the legs, or suffer from a muscle or tendon tear. These factors are associated with various issues that can invoke pain along with developing trigger points along the lower legs. When there are issues in the peroneal muscles, it can lead to muscle weakness in the ankles or “weak ankles,” which causes instability in the body and causes the individual to sprain their ankles. Studies reveal that when the peroneal tendons have a tear in the lower extremities, it can lead to lateral ankle pain that is often missed when examined. However, to that point, if the incision has been left untreated, it can lead to persistent ankle pain, instability, and ankle dysfunction. In “Myofascial Pain and Dysfunction,” written by Dr. Janet G. Travell, M.D., stated that when individuals suffer from weak ankles or have an ankle sprain, active trigger points can cause pain and tenderness to the ankles and cause the person to become unstable. If left untreated, it can cause them to lose balance and have foot drop and ankle fractures to their foot. The book also mentioned that any ruptures in the tendons and muscles might cause lateral compartment syndrome. When there is instability in the ankles, many people resort to using mobility aids like a cane or a walker to be mobile to compensate for the function lost in their feet.
Trigger Point Therapy On The Peroneal Muscles- Video
Do you feel the pain from the bottom of your feet to your ankles? Does it hurt to walk around for a short period? Or have you sprained your ankle, and there is a dull ache when you try to rotate? Some ankle issues are associated with trigger points affecting the peroneal muscles. The peroneal muscles help the lower legs by allowing eversion to the foot and plantarflexion to the ankles. The two muscles that make up the peroneal muscles are the peroneus longus and the peroneus brevis, and they, along with the other tendons and ligaments, help with ankle stability. Since the ankle is a mobile joint, it can succumb to sprains, tears, and instability in the body, allowing trigger points to develop and causing even more issues. The great news is that there are ways to manage trigger points along the peroneal muscles and reduce ankle instability. The video above shows where the peroneal muscles are located on the leg, where the trigger points are, and how to use K-tape to help support the ankle and prevent more injuries on this moveable joint.
Strengthening The Ankles & Managing Trigger Points
Instability in the ankles can be a bummer to many individuals that are on the move, but when it comes to treatment, it can prevent future injuries from re-occurring. Studies reveal that when pain specialists incorporate joint mobilization techniques and dry needling therapy into their patients, it can allow them to reduce the pain and disability to the ankles, thus managing the trigger points along the peroneal muscles. Another way many people can reduce pain in their peroneal muscles is by incorporating stretches and exercises to strengthen their ankles. This allows the peroneal muscles to be loose and gently stretched while slowly strengthening the ankles in a semi-lock position. When people utilize these techniques on their legs and ankles, it can bring mobility and stability back to the body without fear of falling or causing more issues in the ankles.
Conclusion
As one of the most mobile skeletal joints in the lower body, the ankles work together with the legs to provide mobility and stability to the body. The lower legs have various muscles, tendons, and ligaments that travel down and help support the legs and ankles. One of the muscles that provide that support is the peroneal muscle. The peroneal muscles consist of two muscles known as the peroneus longus and peroneus brevis help with eversion to the foot and allow plantarflexion to the ankle. When a person has sprained their ankle, it causes the peroneal muscle to become overstretched and develop trigger points. The great news is that trigger points are treatable, and various treatments can reduce pain in the affected muscle. This allows stability and mobility back to the ankles and improves the body’s functionality.
References
Abd-Rasid, A F, and M Y Bajuri. “Isolated Peroneus Longus Tear – Commonly Missed Diagnosis of Lateral Ankle Pain: A Case Report.” Malaysian Orthopaedic Journal, U.S. National Library of Medicine, July 2020, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7513650/.
Basit, Hajira, et al. “Anatomy, Bony Pelvis and Lower Limb, Foot Peroneus Brevis Muscle.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 8 Feb. 2022, https://www.ncbi.nlm.nih.gov/books/NBK535427/.
Lezak, Bradley, and Matthew Varacallo. “Anatomy, Bony Pelvis and Lower Limb, Calf Peroneus Longus Muscle.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 25 Aug. 2022, https://www.ncbi.nlm.nih.gov/books/NBK546650/.
Salom-Moreno, Jaime, et al. “Trigger Point Dry Needling and Proprioceptive Exercises for the Management of Chronic Ankle Instability: A Randomized Clinical Trial.” Evidence-Based Complementary and Alternative Medicine : ECAM, U.S. National Library of Medicine, 2015, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4430654/.
Travell, J. G., et al. Myofascial Pain and Dysfunction: The Trigger Point Manual: Vol. 2:the Lower Extremities. Williams & Wilkins, 1999.
The legs are crucial for many individuals to move, jump, run, walk, and stand in various locations. The legs involve the thighs, hips, and knees as they work together to provide support and a range of movements for the body. For athletes, the legs allow them to run from one obstacle to another and kick the object to finish the game they are participating. Many individuals require strong leg muscles to keep the body balanced and stabilized from the upper body’s weight. One leg muscle that allows the body to be stabilized is the anterior tibialis muscle. When the legs suffer from various sports injuries or injuries in general, it can lead to issues like shin splints correlated with trigger points that can cause pain to the lower portion of the legs and can affect the body’s stability. Today’s article examines the anterior tibialis muscles, how shin splints are associated with myofascial trigger points, and various methods to treat shin splints. We refer patients to certified providers that incorporate various techniques in the lower body extremities, like lower leg pain therapies correlating to myofascial trigger point pain, to aid many people dealing with pain symptoms along the anterior tibialis muscles, causing shin splints. We encourage and appreciate each patient by referring them to associated medical providers based on their diagnosis, especially when appropriate. We understand that education is an excellent source to asking our providers intricated questions at the patient’s request. Dr. Alex Jimenez, D.C., only utilizes this information as an educational service. Disclaimer
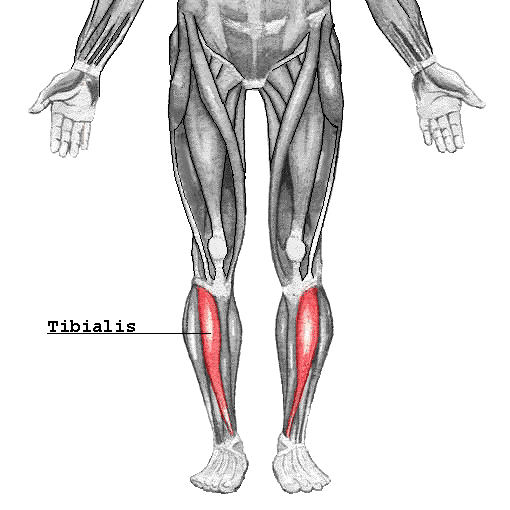
What Is The Tibialis Anterior Muscles?
Have you been dealing with leg pain affecting your ability to move? Do you feel radiating pain going down to your feet? Or does even the smallest amount of pressure sends shooting pain from your knees to your feet? Many of these leg pain issues correlate to myofascial trigger points along the anterior tibialis muscles, mimicking shin splints. Studies reveal that the leg is divided into anterior, lateral, and posterior crural compartments. As one of the largest four muscles in the anterior compartment of the legs, the tibialis anterior is a thick muscle located in the front of the lateral tibia of the legs. The tibialis anterior has the muscle that allows the function to the lower leg and tendons that travel down to the ankle and foot. The anterior tibial muscle plays an important role in the lower leg through dorsiflexion and inversion of the foot. To that point, the anterior tibial muscle plays a key role in energy absorption when walking and maintaining balance.
Shin Splints Associated With Myofascial Trigger Points
Since the anterior tibial muscle plays a key role in energy absorption when it comes to walking and maintaining balance in the body, when the lower leg extremity muscles have been overused, it causes stress on the tibial anterior. It can lead to medial tibial stress syndrome or shin splints. Studies reveal that shin splints affect many athletes, especially runners, by causing pain and discomfort to the tibial anterior. This can cause mobility and balancing issues in the legs and lead to the development of myofascial trigger points in the anterior tibial muscle. Now, how do shin splints and myofascial trigger points correlate with each other?
Dr. Janet G. Travell, M.D., author of “Myofascial Pain and Discomfort: The Trigger Point Manual,” mentioned that one of the chief complaints many people have when experiencing myofascial trigger points would feel muscle weakness of dorsiflexion to the foot when walking. Other complaints include:
Falling
Dragging their feet
Ankle weakness
The book also mentioned that myofascial pain causes referred pain to the anterior tibial muscle, thus mimicking shin splints. The activation from myofascial trigger points causes an overload of the anterior tibial muscle, thus causing various pain issues in the legs and restricting mobility to the muscle itself.
An Overview Of Tibialis Anterior Trigger Points- Video
Have you been dealing with radiating pain from your knees to your feet? Do your legs feel heavy from walking a short distance? Or do your leg muscles feel cramps that hinder your ability to move? These pain-like issues are associated with the anterior tibialis muscle being affected by trigger points. Trigger points or myofascial pain syndrome can affect the worldwide population by affecting a muscle or muscle group in the body that can impair mobility, cause pain-like symptoms, and reduces a person’s overall sense of well-being. Trigger points along the tibialis anterior muscle cause mobility issues and mimic shin splint issues in the legs. All is not lost, however, as there are ways to reduce pain-like symptoms and help manage myofascial trigger points in the anterior tibialis muscle. The video above explains where the trigger points are located in the tibialis anterior through palpitation. By finding the trigger points in the affected muscle, doctors can refer patients to pain specialists who target trigger points and provide treatment to reduce the pain.
Various Methods Of Treating Shin Splints
There are various methods to treat the tibialis anterior when treating shin splints associated with trigger points. Studies reveal that one of the multiple ways to reduce shin splints is to strengthen the core hip muscles, improve running mechanics, and prevent lower-extremity overuse injuries. Muscle strength training allows the other muscles from the abdominals, gluteal, and hips to be stronger and reduce strain on the anterior tibialis muscles. Another method that many individuals should consider is to wear the appropriate footwear. Wearing the proper footwear can reduce the shock absorption to the feet and reduce the overloading forces on the anterior tibialis. These are two methods to manage trigger points and prevent shin splints from re-occurring in the legs.
Conclusion
As one of the four leg muscles, the anterior tibialis is a large muscle located in front of the lateral tibia and travels down to the ankles and foot. This muscle plays an important role in the legs as it allows dorsiflexion and inversion of the foot while also playing a key role in energy absorption when walking and maintaining balance. When the anterior tibialis becomes overused, it can develop trigger points, which invoke shin splints in the legs. When the legs suffer from shin splints associated with trigger points, it can cause pain in the lower leg extremities and cause the body to become unstable. However, various methods can take the load off the tibialis anterior and help improve the body’s stability, allowing the individual to walk without feeling pain traveling up from their feet.
References
Deshmukh, Nikita S, and Pratik Phansopkar. “Medial Tibial Stress Syndrome: A Review Article.” Cureus, U.S. National Library of Medicine, 7 July 2022, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9356648/.
Galbraith, R Michael, and Mark E Lavallee. “Medial Tibial Stress Syndrome: Conservative Treatment Options.” Current Reviews in Musculoskeletal Medicine, U.S. National Library of Medicine, 7 Oct. 2009, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2848339/.
Juneja, Pallavi, and John B Hubbard. “Anatomy, Bony Pelvis and Lower Limb, Tibialis Anterior Muscles.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 29 Aug. 2022, https://www.ncbi.nlm.nih.gov/books/NBK513304/.
Travell, J. G., et al. Myofascial Pain and Dysfunction: The Trigger Point Manual: Vol. 2:the Lower Extremities. Williams & Wilkins, 1999.
Zielinska, Nicol, et al. “Anatomical Variations of the Tibialis Anterior Tendon Insertion: An Updated and Comprehensive Review.” Journal of Clinical Medicine, U.S. National Library of Medicine, 19 Aug. 2021, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8396864/.
As one of the stabilizers for the body, the knees are located between the thighs and legs, allowing flexion and extension. The knees help the hips by supporting the upper body’s weight and allowing the legs to move from one place to another without feeling pain. The knee has various muscles and ligaments surrounding the knee joint, allowing the leg to be bent when active. One of the muscles is located behind the knee, known as the popliteus, and supports the legs. However, minor injuries or actions can affect the knees causing the joint to be in a “lock” position and develop myofascial trigger points that can induce muscle spasms in the knees. Today’s article focuses on the popliteus muscle, how knee pain is associated with trigger points, and how to manage knee pain through various treatments. We refer patients to certified providers that incorporate multiple methods in the lower body extremities, like knee pain treatments correlating to myofascial trigger points, to aid many people dealing with pain symptoms along the popliteus muscles. We encourage and appreciate each patient by referring them to associated medical providers based on their diagnosis, especially when appropriate. We understand that education is an excellent source to asking our providers intricated questions at the patient’s request. Dr. Alex Jimenez, D.C., only utilizes this information as an educational service. Disclaimer
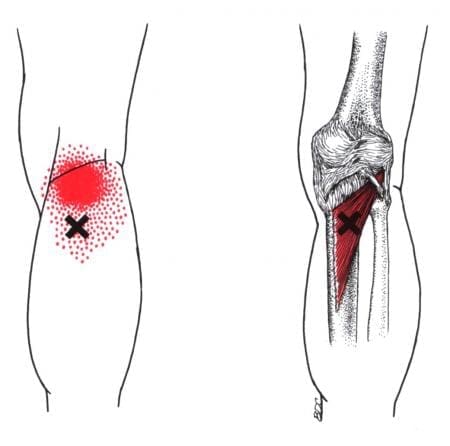
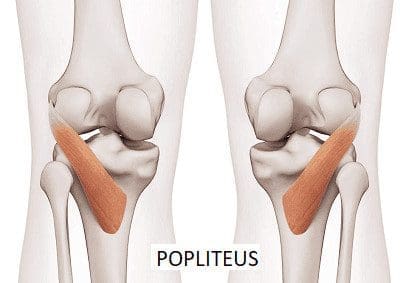
What Is The Popliteus Muscle?
Have you been dealing with pain behind your knees? Do you have issues bending your knees when climbing up or down the stairs? Or do your back knee muscles start to twitch uncontrollably, causing muscle spasms? Many knee issues correlate with various factors that can affect the popliteus muscle and develop trigger points. The popliteus is a small muscle with a very important job as it is a major stabilizing muscle to the knees. The popliteus muscle originates from the lateral side of the femur and inserts itself into the posterior surface of the tibia. Some attachments are between the popliteus and lateral meniscus, allowing the knees to be in motion and providing flexion without pain and entrapment. Additional studies reveal that when a person exercises, the popliteus’s basic function helps bring about and maintain internal rotation of the tibia on the femur. The popliteus also helps prevent the foot from external rotation and allows the individual to stand correctly. However, injuries to the knee could overstretch the popliteus muscle and cause mobility issues to the knee flexion.
Knee Pain Associated With Trigger Points
When dealing with knee pain, it could often be a joint disorder like osteoarthritis or a musculoskeletal condition like sciatica pain associated with the knee. These issues could be due to normal factors like constantly sitting down or bending down to lift heavy objects that cause the knees to buckle. However, when the popliteus muscle has been continuously overused from being bent, it can form tiny nodules known as trigger points to cause knee pain. Studies reveal that trigger points on the muscles surrounding the knee are often ignored during a clinical diagnosis. Trigger points cause referred pain to the surrounding muscles, accompanied by various sensory sensations like heaviness, tingling, and hypersensitivity to the popliteus muscle. In “Myofascial Pain and Dysfunction,” written by Dr. Travell, M.D. stated that one of the chief complaints that many patients often talk to their doctors about is the pain they feel in the back of their knees when they are in a crouch position. The book also states when normal actions like running or twisting have overloaded the popliteus muscle, it can cause trauma or strain to the popliteus muscle and tear the posterior cruciate ligament to the knees.
How To Find Trigger Points In The Popliteus- Video
Have you been having knee issues that make walking difficult for a long period? Do you feel like your knees are locking up constantly? What about feeling unstable when standing or carrying objects around? These issues that affect the knees are associated with trigger points along the popliteus muscles. The popliteus muscle is small, located at the back of the knees, and assists with knee flexion. When the popliteus muscle becomes overused, it can cause trigger points to form and cause knee issues. Studies reveal that various issues, like tendon injuries, are associated with repetitive mechanical stresses that can cause degenerative knee lesions. Any trauma or muscle strain can affect the knee’s function of flexing and bending without pain for trigger points to form along the popliteus muscles. The video above focuses on the popliteus muscle, where the trigger points are located, and where the referred pain patterns are situated in the knees. On the bright side, all is not lost, as various treatments offer ways to manage knee pain associated with trigger points.
Managing Knee Pain Through Various Treatments
When it comes to knee pain, many individuals will apply an ice or heat compress to allow the surrounding muscles to relax while reducing the pain and swelling. Other individuals use over-the-counter medicines to eliminate the pain for a few hours. While these work at managing knee pain, various treatments target trigger points and can help improve flexion mobility back to the knees. Studies reveal that muscle stretching on the popliteus muscle contributes to joint position sense to knee joint stability and function. Stretching the popliteus muscles can reduce the pain in the back of the knee while elongating the muscle fibers to manage trigger points from forming again. Other treatments that people can do to avoid trigger points from returning is to avoid walking or running in a lateral sloped area to prevent the knees from locking up. Incorporating these treatments to prevent knee issues and allow the knee to function properly.
Conclusion
The knees are one of the stabilizers in the body that are located between the thighs and legs, allowing flexion and extension. As a small muscle located in the back of the knees, the popliteus stabilizes the knees and enables them to be in motion without pain. However, when the popliteus muscle becomes overstretched and overused, it can develop trigger points in the popliteus that invoke referred pain to the surrounding muscles and cause the knees to lock up. To that point, it causes the body to be unstable and mimics knee pain issues. Fortunately, trigger points are treatable through various treatments that help relieve the pain and reduce the trigger points from returning. When these treatments are utilized on the knees, the surrounding muscles regain flexion mobility in the lower body.
References
English, S, and D Perret. “Posterior Knee Pain.” Current Reviews in Musculoskeletal Medicine, U.S. National Library of Medicine, 12 June 2010, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2941578/.
Ghaffarinejad, Farahnaz, et al. “Effect of Static Stretching of Muscles Surrounding the Knee on Knee Joint Position Sense.” British Journal of Sports Medicine, U.S. National Library of Medicine, Oct. 2007, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2465159/.
Hyland, Scott, and Matthew Varacallo. “Anatomy, Bony Pelvis and Lower Limb, Popliteus Muscle.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 6 June 2022, https://www.ncbi.nlm.nih.gov/books/NBK526084/.
Mann, R A, and J L Hagy. “The Popliteus Muscle.” The Journal of Bone and Joint Surgery. American Volume, U.S. National Library of Medicine, Oct. 1977, https://pubmed.ncbi.nlm.nih.gov/908724/.
Sánchez Romero, Eleuterio A, et al. “Prevalence of Myofascial Trigger Points in Patients with Mild to Moderate Painful Knee Osteoarthritis: A Secondary Analysis.” Journal of Clinical Medicine, U.S. National Library of Medicine, 7 Aug. 2020, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7464556/.
Travell, J. G., et al. Myofascial Pain and Dysfunction: The Trigger Point Manual: Vol. 2:the Lower Extremities. Williams & Wilkins, 1999.
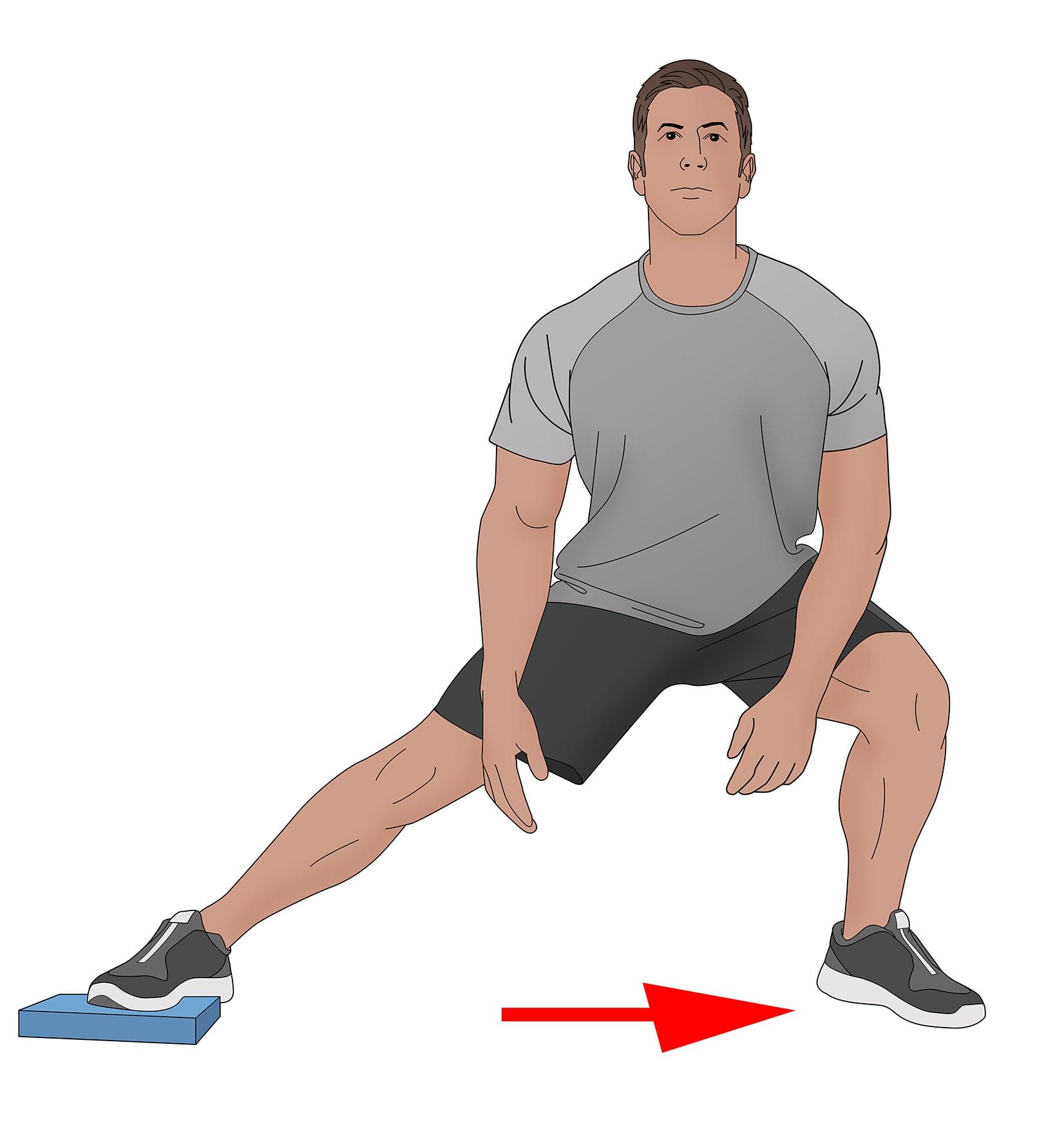
Many individuals utilize their lower muscles to move around and stay active as each muscle does its job and allows mobility to the hips and thighs. In sports, the thigh muscles are utilized constantly to extend the legs and bend the knees, allowing a powerful force to win any sports competition. At the same time, various sports injuries can occur to the hips, thighs, and legs and can affect the muscles causing pain and discomfort to the lower extremities. A hamstring injury is one of the most common injuries that can affect the thighs, which can cause many athletes to be taken out of their favorite sport to recover from the injury. Today’s article looks at the hamstring muscle, how trigger points correlate with a hamstring strain, and how various stretches can reduce muscle strain on the hamstrings. We refer patients to certified providers who incorporate multiple methods in the lower body extremities, like upper thigh and hip pain treatments correlating to myofascial trigger point pain, to aid individuals dealing with pain symptoms along the hamstring muscles. We encourage and appreciate patients by referring them to associated medical providers based on their diagnosis, especially when appropriate. We understand that education is an excellent solution to asking our providers complex questions at the patient’s request. Dr. Jimenez, D.C., utilizes this information as an educational service only. Disclaimer
What Are The Hamstring Muscles?
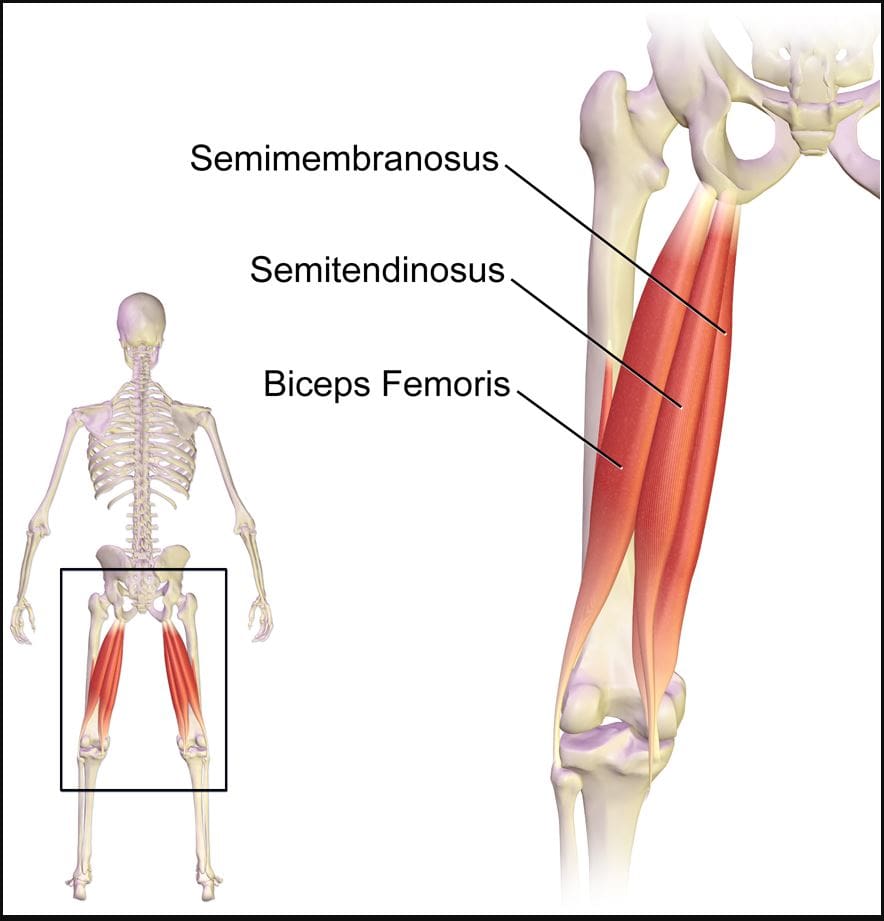
Do you experience pain in the back of your upper thigh? When walking from one place to another, do you hear a popping sound in the back of your thigh? Or are you dealing with muscle tenderness in the back of your upper thigh? Many of these symptoms correlate with issues affecting the hamstrings causing trigger points to affect the upper thighs. As one of the most complex muscles comprising three muscles (semitendinosus, semimembranosus, biceps femoris), the hamstrings play a crucial part in daily activities. From simple actions like standing to explosive movements like sprinting or jumping, the hamstrings are known as posterior thigh muscles that begin from the pelvis and run behind the femur bone and cross the femoroacetabular and tibiofemoral joints. The hamstring muscles in the body play a prominent role in hip extension and is a dynamic stabilizer of the knee joint. To that point, the hamstring muscles are the most susceptible muscle that succumbs to injuries that can lead to disability in the legs and affect daily activities.
Hamstring Strain & Trigger Points
Since the hamstrings are the most susceptible muscles that can succumb to injuries, it takes a while for the muscle to heal, depending on the severity of the damage. Studies reveal that the hamstrings can occur injuries when a person is running or sprinting due to their anatomic arrangement, which causes the muscles to strain. To that point, depending on how much force has impacted the hamstrings, the injuries can lead to 3 of the following:
Grade 1: Mild pain or swelling (no loss of function)
Grade 2: Identifiable partial tissue disruption with moderate pain and swelling (minimal loss of function)
Grade 3: Complete disruption of the tissue with severe pain and swelling (total loss of function)
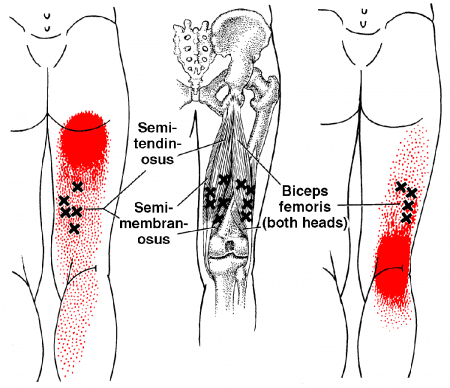
The pain that patients experience can be painful when walking, causing them to limp. In “Myofascial Pain and Dysfunction,” written by Dr. Janet G. Travell, M.D., stated that when patients are dealing with pain in their hamstrings, it could potentially be associated with trigger points along the three muscles, causing pain and disability in the upper thighs. The book also mentioned that when trigger points affect the hamstrings, it can lead to muscle inhibition, compromising hip stability. Another issue that trigger points associated with hamstring strain causes in the body are that when individuals are sitting down are likely to experience posterior pain in the buttock, upper thighs, and back of the knees. Luckily, there are various ways to reduce the pain along the hamstring muscles.
Trigger Point Of The Week: Hamstrings- Video
Have you dealt with pain along the back of your upper thighs? Does it feel uncomfortable when you are sitting down? Or do your hamstrings ache or feel tight after running for a long period? People dealing with issues in their hamstrings could be dealing with muscle strain associated with trigger points. The hamstring muscles play a vital role in the body as it allows the individual to walk, run, bend the knees and even extend the legs. The hamstring muscles are also the most susceptible to injury, causing disability to the legs. Studies reveal that trigger points associated with the hamstring muscles can lead to soreness or irritability in the muscle fibers that may interfere with the biomechanics and normal functioning of the lower limbs. The video above explains where the hamstrings are located and how the trigger points can cause referred pain to the hamstrings. To that point, trigger points can affect a person’s ability to walk and affect the surrounding muscles in the lower body while mimicking other chronic conditions.
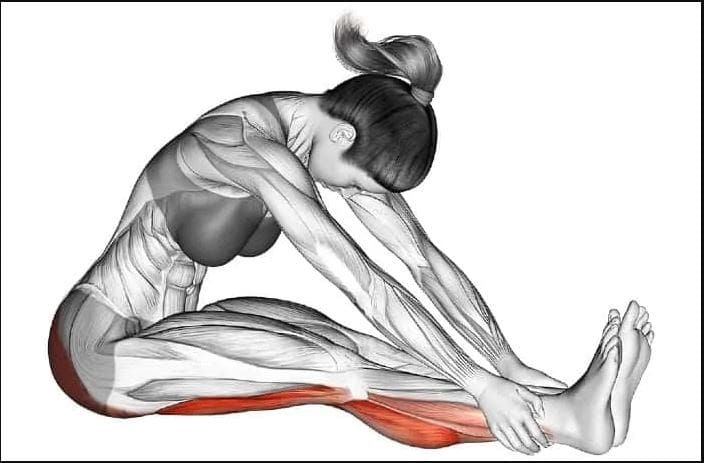
Various Stretches To Reduce Muscle Strain On The Hamstrings
When the hamstrings become injured, the healing rate usually depends on how severe the injury is in the hamstrings. If a hamstring injury is mild, the tears or strains can heal within about three to eight weeks, and if the hamstring injury is severe, the tears or strains could be long as three months. When the hamstrings are tense and on the verge of tearing, many people should stop overusing the muscle. Various stretches can reduce muscle strain on the hamstrings and relieve tension from the hamstrings to allow mobility back to the legs. Studies reveal that manual ischemic compression on the upper thigh muscles can significantly reduce pain in the lower limbs. This allows the individual to manage the trigger points associated with the hamstrings and reduce the chances of them re-occurring in the legs.
Conclusion
As the most important muscle in the lower body extremities, the hamstrings play a crucial part in the body as they allow the individual to walk, run, and stand without feeling pain. However, even though they are important muscles, they are susceptible to injuries. When the hamstrings become injured, the recovery process varies depending on the severity and can develop trigger points along the muscle fibers. To that point, it causes referred pain along the upper thigh muscle and affects a person’s ability to walk. Fortunately, incorporating various stretches to the hamstrings can alleviate the pain and reduce the trigger points from re-occurring in the muscle. This allows mobility back to the legs, and many individuals can resume their daily activities.
References
Esparza, Danilo, et al. “Effects of Local Ischemic Compression on Upper Limb Latent Myofascial Trigger Points: A Study of Subjective Pain and Linear Motor Performance.” Rehabilitation Research and Practice, Hindawi, 4 Mar. 2019, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6425406/.
Poudel, Bikash, and Shivlal Pandey. “Hamstring Injury – Statpearls – NCBI Bookshelf.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 28 Aug. 2022, https://www.ncbi.nlm.nih.gov/books/NBK558936/.
Rodgers, Cooper D, and Avaias Raja. “Anatomy, Bony Pelvis and Lower Limb, Hamstring Muscle.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 29 Jan. 2022, https://www.ncbi.nlm.nih.gov/books/NBK546688/.
Thummar, Ravindra C, et al. “Association between Trigger Points in Hamstring, Posterior Leg, Foot Muscles and Plantar Fasciopathy: A Cross- Sectional Study.” Journal of Bodywork and Movement Therapies, U.S. National Library of Medicine, 7 Aug. 2020, https://pubmed.ncbi.nlm.nih.gov/33218537/.
Travell, J. G., et al. Myofascial Pain and Dysfunction: The Trigger Point Manual: Vol. 2:the Lower Extremities. Williams & Wilkins, 1999.
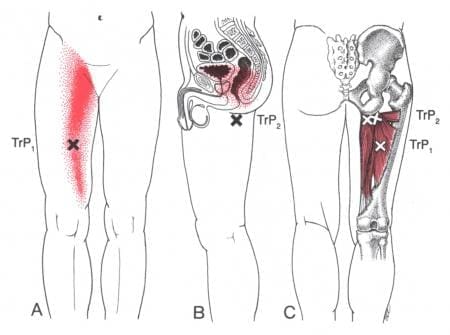
The hips and thighs have a working relationship as their jobs are to maintain stability for the legs and pelvis while supporting the upper body’s weight. These two body groups have various muscles, tendons, and nerves that have specific jobs that allow mobility to the lower body. Many athletes in multiple sports events use their thighs to exert a huge amount of power to be the best. This is due to the adductor muscles in the thighs that allow the athlete to win the event. These adductor muscles are voluminous in size and can become overstretched if the muscles have been worked out too much or injuries have caused dysfunction in the surrounding muscles, causing mobility issues. To that point, the adductor muscles will develop myofascial trigger points and cause hip and thigh pain. Today’s article looks at the two adductor muscles (Longus and Magnus), how myofascial trigger points affect the adductor muscles, and available treatments to manage hip adductor trigger points. We refer patients to certified providers who incorporate multiple methods in the lower body extremities, like thigh and hip pain treatments correlating to myofascial trigger point pain, to aid individuals dealing with pain symptoms along the adductor muscles. We encourage and appreciate patients by referring them to associated medical providers based on their diagnosis, especially when appropriate. We understand that education is an excellent solution to asking our providers complex questions at the patient’s request. Dr. Jimenez, D.C., utilizes this information as an educational service only. Disclaimer
Adductor Longus & Adductor Magnus
Have you been dealing with groin pain located near your thighs? Do you feel muscle tenderness or stiffness when stretching your inner thighs? Or have you been feeling unstable in your hips or thighs when walking? Many individuals, especially athletes and older adults, could be experiencing myofascial trigger points associated with groin pain along their adductor muscles. The thighs contain several muscles and functions that allow many people to bend and extend their knees and hips. The adductor muscles allow the legs to move inward toward one another. The adductor muscles have five muscles: magnus, brevi, longus, pectineus, and gracilis. These muscles enable functionality to the thighs and hips, and we will look at two adductor muscles in the inner thighs. The long adductor muscle is a large, fan-shaped muscle that starts from the superior aspect of the pubis bone and travels down to connect at the thigh bone. Studies reveal that the adductor longus is a long and thin muscle with many actions for the thighs, including external/lateral rotation and thigh flexion.
Now the adductor Magnus is a large triangular-shaped muscle of the inner thighs that are important for thigh and hip function and stabilizing the pelvis. Studies reveal that even though the adductor Magnus is a large muscle in the inner thighs, its primary function is to allow the thigh to move in a larger range of motion without any pain inflicted on the thigh muscles. However, the adductor muscle can succumb to various issues affecting the thighs and groin regions of the body that can be overstretched and strain the body.
Myofascial Trigger Points Affecting The Adductor Muscles
Groin pain is a multi-factorial pain issue that affects the lower limbs, and its often due to muscle strain in the inner thigh muscles. This pain increases during vigorous activities and when there is a sudden twist in the hips. When the adductor muscles suddenly change in motion when the body is active, they can be overstretched and correlate to myofascial trigger points that can affect the inner thigh and groin regions. According to “Myofascial Pain and Dysfunction,” by Dr. Travell, M.D., patients with active myofascial trigger points in the two adductor muscles (Longus and Magnus) would become frequently aware of the pain in their groin and medial thigh. When the adductor muscles have myofascial trigger points in the inner thigh, diagnosing is difficult since the individual thinks they are suffering from groin pain when the pain is in their inner thighs. To that point, studies reveal that many individuals participating in various sports would suffer from groin pain due to myofascial trigger points affecting the adductor muscles. Luckily, there are multiple treatments to reduce the pain in the adductor muscles.
Hip Adductors: Trigger Point Anatomy- Video
Have you been dealing with groin pain when you are walking? What about experiencing unquestionable thigh pain that affects your daily activities? Or does stretching your inner thigh muscles seem difficult, causing muscle tenderness? Many of these symptoms correlate with groin pain associated with myofascial trigger points affecting the adductor muscles in the inner thighs. The adductor muscles allow mobility function to the thighs and enable the hips to have a wide range of motion. When the adductor muscles are overstretched due to a sudden change of hip rotation or injury has occurred on the thighs can lead to referred pain in the groin and inner thighs and develop myofascial trigger points. The video above shows where the trigger points are located in the hip adductor muscles. The video also explains where the pain is localized in the adductor muscles and the symptoms it produces that can affect the lower body extremities. Fortunately, even though diagnosing myofascial trigger points are a bit challenging, available treatments can manage trigger points along the hip adductors.
Available Treatments To Manage Hip Adductor Trigger Points
When myofascial trigger points affect the hip adductor muscles, many individuals complain about stiffness in their inner thighs and how they feel miserable when they don’t have mobility from their thighs and hips. As stated earlier, trigger points are a bit challenging when diagnosed, but they are treatable when doctors examine patients dealing with myofascial pain in their hips and thigh muscles. Once the diagnosis is complete, doctors work with pain specialists who can locate the trigger points and devise a treatment plan to relieve the pain. Available treatments like trigger point injections can minimize the pain and reduce the chances of trigger points returning. Other available therapies like exercising or stretching, especially for the hips and thighs. Specific exercises for the hips and thigh muscles can help strengthen the adductor muscles from suffering pain and can help reduce the pain symptoms. Another treatment is applying moist heat on the hip adductor muscles to release the tension from the tight muscles and allow mobility back to the hip adductors.
Conclusion
The adductor muscles work with the hips and thighs to allow a wide range of motions and extension to the knees and hips. The hips and the thighs allow stability to the lower body and support the weight to the upper body. When injuries or sudden changes start to affect the adductor muscles, it can lead to symptoms of groin pain associated with myofascial trigger points. Myofascial trigger points produce tiny nodules in the affected muscle that causes referred pain to the muscle group. When this happens, it causes the body to be dysfunctional and can affect a person’s mobility to function in the world. Luckily myofascial trigger points are treatable through various techniques and treatments that can reduce the chances of trigger points from re-occurring in the body.
References
Jeno, Susan H, and Gary S Schindler. “Anatomy, Bony Pelvis and Lower Limb, Thigh Adductor Magnus Muscle.” In: StatPearls [Internet]. Treasure Island (FL), StatPearls Publishing, 1 Aug. 2022, https://www.ncbi.nlm.nih.gov/books/NBK534842/.
Sedaghati, Parisa, et al. “Review of Sport-Induced Groin Injuries.” Trauma Monthly, Kowsar, Dec. 2013, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3864393/.
Simons, D. G., and L. S. Simons. Myofascial Pain and Dysfunction: The Trigger Point Manual: Vol. 2:the Lower Extremities. Williams & Wilkins, 1999.
Takizawa, M, et al. “Why Adductor Magnus Muscle Is Large: The Function Based on Muscle Morphology in Cadavers.” Scandinavian Journal of Medicine & Science in Sports, U.S. National Library of Medicine, 27 Apr. 2012, https://pubmed.ncbi.nlm.nih.gov/22537037/.
van de Kimmenade, R J L L, et al. “A Rare Case of Adductor Longus Muscle Rupture.” Case Reports in Orthopedics, Hindawi Publishing Corporation, 2015, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4397006/.
IFM's Find A Practitioner tool is the largest referral network in Functional Medicine, created to help patients locate Functional Medicine practitioners anywhere in the world. IFM Certified Practitioners are listed first in the search results, given their extensive education in Functional Medicine