The fasting mimicking diet is an alternative to fasting. However, it can have several benefits for your overall health and wellness. We will discuss everything you need to know about the regimen. The article below describes how to do it, its benefits, and how it’s different from normal fasting. The benefits of the fast mimicking diet will have you wanting to try it for yourself.
What is the Fasting Mimicking Diet?
The fasting mimicking diet is a type of modified fasting. The regimen produces the same benefits of fasting by eating small amounts of food. The fast mimicking diet generally lasts about five days and it includes a healthy protocol of carbohydrates, proteins, and fats.
Calories are also maintained at approximately 40 percent of the average calorie intake. This permits the human body to remain nourished without the stress of normal fasting. Calorie restriction can cause health issues, however, the fast mimicking diet is safe and effective. Below, we will discuss just how much the fast mimicking diet differs from traditional fasting.
Traditional Fasting Vs Fast Mimicking Diet
The fasting mimicking diet is always compared to intermittent fasting. There are many myths about these types of modified fasts. Some claim that our muscles waste away while others claim that they change our metabolism, and that it’s downright unhealthy.
The health issues discussed above may be true for a person who’s actually restricting their calorie intake. Some types of fasting may cause metabolic damage which may not be recommended for people with underlying health conditions. However, the fast mimicking diet gives you all the advantages of fasting without the side effects. Below are the benefits of the fast mimicking diet.
Benefits of the Fast Mimicking Diet
The benefits of the fast mimicking diet are essentially the same as those of regular fasting. The benefits are listed below.
The fast mimicking diet “tricks” the human body into feeling as though it’s fasting. Now that we have discussed what this alternate form of fasting is and why it is worth doing, the following advice will demonstrate how to do the diet itself.
How to do the Fast Mimicking Diet
Research studies have found that the best results for the fasting mimicking diet occur in about five days or when your glucose ketone index drops below 1.0. Doing this regimen anywhere between 3 to 7 days is also beneficial. The regimen should also be repeated every month to fully experience its benefits, unless otherwise instructed by a healthcare professional.
If you’re interested in monitoring your fasting outcomes, you should consider quantifying specific biomarkers. This could be measured through lab tests before and after following the fasting mimicking diet. Measuring blood glucose, ketones, and weight changes every day can also be helpful to determine your biomarkers. You might also want to set up your environment by:
Telling friends and family about what you are doing and asking them for their support.
Eliminating any snack foods at home or work that might interrupt your regimen.
Giving yourself more time to sleep, as you will probably be more exhausted than usual.
Planning for exercise and physical activity every day. But keep away from intense workouts during this time.
Now that we discussed how you can do the diet, let’s discuss the basics of the fast mimicking diet.
The fasting mimicking diet provides the same great benefits of fasting while still providing your body with some nourishment. If you are following this regimen, make sure that you maintain a low-calorie intake and utilize appropriate supplements to achieve ketosis without experiencing health issues. Set up your environment for the diet. And if you decide to blend the ketogenic diet with this alternate form of fasting to get into ketosis faster, you can achieve the maximum advantages out of the two regimens. Be sure to consult a healthcare professional before following the fasting mimicking diet. Dr. Alex Jimenez D.C., C.C.S.T. Insight
Fasting Mimicking Diet Basics
Some people today might eat a slightly higher amount of calories the first day as they ease into the fasting mimicking diet. They might then decrease their total caloric intake. You also want to make sure you eat smaller amounts of foods which are easy to digest.
ProLon� offers a pre-packaged box which contains all five days’ worth of meals for you to do the diet. The meals are all plant-based. One day, by way of instance, offers tea and a nut bar for breakfast, a small portion of vegetable soup and a few kale crackers for lunch, several olives in the afternoon, and finally another small portion of vegetable soup for dinner.
You can also do the fasting mimicking diet without the need for a pre-packaged box like ProLon�. Simply follow the right proportions and plan out how you will space them out every day. Macros for the fast mimicking diet are 34 percent carbohydrates, 10 percent protein, and 56 percent fat for the very first day and 47 percent carbohydrates, 9 percent protein, and 44 percent fat to the rest days.
A cup of black tea and coffee every day are generally allowed. Just make sure they don’t contain any added sugars or oils. Remember that people with health issues should consult a healthcare professional prior to doing the fast mimicking diet in your own home.
Foods
Dr. Anthony Gusting followed a four-day ketogenic fasting mimicking diet. Every day, he consumed different amounts of bone broth, coconut milk, coconut oil, BCAAs, and exogenous ketones. Avocados and grass-fed butter can also be included in the fast mimicking diet. This is a great way to combine the ketogenic diet with the fasting mimicking�diet to benefit from the two regimens.
Supplements
Taking nutritional supplements can also make the fasting mimicking diet easier by providing enough nutrition. These may include:
Electrolytes like magnesium and salt to replenish any lost during water loss
Grass-fed liver tablets to provide micronutrient support
Branch chain amino acids, or BCAAs, to help prevent loss of lean tissue
Greens powder to provide micronutrients
Algal oil or cod liver oil for omega-3s
You may also take exogenous ketones to achieve ketosis through the keto diet. The fast mimicking diet can also help you achieve ketosis before following a ketogenic diet. Below, we will discuss how the fast mimicking diet promotes ketosis.
Ketosis and the Fast Mimicking Diet
The fast mimicking diet is an excellent way to prepare you for the ketogenic diet. This is because it allows you to get into ketosis. Additionally, eating keto foods makes it possible to remain in ketosis throughout the regimen. To follow a ketogenic fasting mimicking diet you must maintain your macros over the suitable range of 5 to 10 percent of carbohydrates, 20 to 25percent of proteins, and 70 to 80 percent of fats. If you’re unsure about whether you’re properly maintaining your macros, always choose something with more fat.
The scope of our information is limited to chiropractic and spinal health issues as well as functional medicine topics and discussions. To further discuss the subject matter, please feel free to ask Dr. Alex Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topic Discussion:�Acute Back Pain
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief. �
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download
* All the above XYMOGEN policies remain strictly in force.
Functional Medicine Doctor Explains Women’s Hormones
We discussed the basics for men�s hormones. Now let�s discuss the basics for women�s hormones. Unfortunately, the effects of our diet and our environment become more obvious in the anatomy and biochemistry of women. These may frequently manifest as hormone imbalances and they can greatly affect their quality of life. Mood disorders have become an epidemic.
By way of instance, depression affects 20 percent of women, about twice as much in women than in men. And premenstrual syndrome, or PMS, affects between 60 and 75 percent of women in the United States. Infertility is also an epidemic which affects more than one in seven couples and it is generally managed by reproductive endocrinologists through invasive procedures, hormone treatments, and in vitro fertilization, or IVF, often without even evaluating what is the cause of the reproductive health issues. I�ve helped many women improve their hormone imbalances through the basic principles of functional medicine.
Now, these are only several of the reasons why we need to determine the source of hormonal imbalances in women. And I believe this knowledge can help women find the answers they need to improve their overall health and wellness. Most importantly, you need to learn to listen to what your body is telling you. After all, the human body is one of the best doctors.
Taking Control of Women’s Hormones
Women�s hormones are much more complex than men�s hormones because they�re constantly changing based on their cycle as well as on their stage of life. If you�re experiencing mood swings, irregular cycles, menstrual pain, heavy bleeding, infertility, weight gain, and brain fog, functional medicine can help improve your symptoms by balancing your hormones.
So, let�s discuss the differences between pre-menopausal women and post-menopausal women. Pre-menopausal women should experience regular cycles every 28 days that last two or three days without a lot of pain, not too heavy bleeding, and no PMS. However, most women don�t experience regular cycles. A proper nutrition consisting of low sugar and starch, high fat, and more fiber can help correct abnormal cycles. A plant-rich diet can also help improve abnormal cycles. Caffeine and alcohol consumption can even cause hormone imbalances in women. Proper nutrition, including taking supplements like magnesium, B vitamins and fish oil, exercise, sleep, and stress management can help regulate your hormones. This is generally enough to help most women.
A doctor or functional medicine practitioner can also help balance your hormones. Because women�s hormones fluctuate throughout their cycle, progesterone and estrogen levels may be different for each woman. Also, depending on the hormonal health issues, patients may need to run their hormone lab panel tests on either day three or day 21 of their cycle for best results.
Furthermore, we will also need to evaluate your ratio of hormones, like that of estrogen to progesterone, because these can be the cause of numerous symptoms for many women. Estrogen dominance is one of the most common problems associated with a woman�s hormone ratio. Progesterone levels in women should be at their highest during the last half of their cycle. The hormone ratio between progesterone and estrogen should be 10 to one. However, if the human body is not producing the required amounts of progesterone, symptoms of estrogen dominance may begin to manifest, regardless if the human body�s estrogen levels themselves are low. Symptoms of estrogen dominance can include: anxiety, heavy bleeding, PMS, breast tenderness, shorter cycles or spotting between cycles, infertility, fluid retention, weight gain, and sleeping problems.
FSH is produced by the brain to help the follicles and the ovaries prepare to release an egg. LH is another hormone produced by the brain which triggers the release of an egg into the uterus so that conception can occur. LH also helps produce progesterone during the second half of your cycle, which may be another reason why many women have low progesterone levels.
These hormones are fundamental to look at if you�re trying to have a baby. The elevated hormone ratio between LH and FSH can demonstrate the presence of a common health issue known as PCOS or polycystic ovarian syndrome. PCOS is actually not an ovarian health issue. As a matter of fact, it�s a common problem associated with a poor diet and insulin resistance. An increased consumption of sugar and starch can cause irregular cycles, heavy bleeding, acne, hair loss, and infertility.
As for post-menopausal women, hormone blood panel tests are just as important and we generally don�t need to worry about having to evaluate them on a specific day of the month. Also, when we test a woman who�s in their perimenopause, their hormones may be tremendously unstable. Therefore, it�s ultimately essential to diagnose a woman�s symptoms to help treat the source of the health issue.
We previously discussed the importance of testosterone in men. However, testosterone is also important in women. Many women visit numerous doctors after experiencing low energy levels as well as a decreased sex drive. Most doctors will associate these symptoms with aging or they may even tell them it�s all in their head and simply prescribe them some Prozac. But if we were to run a blood panel test on them, their testosterone levels would often come back undetectable. It�s no wonder why women don�t feel like themselves after they�ve lost their libido and their vitality.
Total testosterone levels in women should be between 60 and 80 while free testosterone levels should be over 0.5. Testosterone is fundamental towards maintaining lean muscle mass and optimizing energy. Testosterone is also important for clear brain function.
Now women, if you have low testosterone levels, this may be causing you to experience a reduced sex drive or it may even be making you feel fatigue. However, this isn�t always the cause of these symptoms. That�s why it�s so essential for women to test their sex hormone levels. Testosterone is what is known as an androgen, or a male hormone, but it�s also found in women. Other androgens that help contribute to male characteristics include androstenedione, dihydrotestosterone, or DHT, and DHEA. In polycystic ovarian syndrome, or PCOS, women will commonly develop elevated levels of any of these hormones. Women who consume a lot of sugar and starch may also develop acne, hair loss or they may even grow facial hair. All of these are symptoms of too much testosterone in women.
PCOS affects approximately 8 to 12 percent of women. As previously mentioned, this health issue is a metabolic problem caused by poor nutrition which ultimately affects the human body�s insulin levels. It can also affect other hormones, such as the androgens we previously discussed. When women develop insulin resistance, the production of male sex hormones can increase. Other tests are important when PCOS is caused by FSH and LH hormones.
FSH generally triggers ovulation. However, if a woman�s FSH is too low due to PCOS, ovulation, and therefore, conception can�t occur. This is why women with PCOS are also commonly diagnosed with infertility. And the key is in a woman�s LH to FSH ratio. Increased levels of LH can stimulate androgens, such as testosterone, and decreased levels of FSH can stimulate the follicles and estrogen. Furthermore, facial hair or thinning of the head hair, irregular menstrual cycles, heavy bleeding, and weight gain in women can be symptoms of PCOS. Although PCOS is believed to be a health issue which exclusively affects overweight women, we�re starting to see an increase of women with healthy weights develop PCOS.
Because functional medicine focuses on finding the source of the health issue, if we see cysts on your ovaries or if any other lab tests indicate the presence of PCOS, we won�t simply stop there. And most often, we just have to look back at the patient�s diet. The high consumption of sugar and starch causes a tremendous metabolic disturbance which can cause a variety of other health issues. We will discuss these various metabolic conditions in another article.
Hormones are fundamental to women’s health. Female sex hormones, including estrogen and progesterone, influence a woman’s mood, menstruation, pregnancy, menopause, and more. A variety of other hormones can also affect other aspects of a woman’s health. One of the most common health issues associated with hormonal imbalances in women is known as polycystic ovarian syndrome or PCOS. Women can be tremendously affected by hormone fluctuations. It’s essential for women to seek help from a doctor to find out if their symptoms are caused by hormone imbalances. Functional medicine can also help regulate hormones. Dr. Alex Jimenez D.C., C.C.S.T. Insight
Understanding Women’s Hormones
We measure hormonal imbalances through simple blood tests and we can also measure hormones through urine tests. Tests known as the �DUTCH� and the �Essential Estrogens� are provided by Genova to determine hormone metabolites. These can also determine the downstream breakdown products of hormones in order to help demonstrate what�s happening with your hormone metabolism.
Now, let me explain what are some of the most important things doctors or functional medicine practitioners look at when ordering a DUTCH test or an Essential Estrogens test. Hormone metabolism tests demonstrate your absolute hormone levels as well as which types of metabolites are being triggered. And this is what we utilize to look at your estrogen levels, androgen levels, and progesterone levels, as well as your cortisol levels, which we will discuss later. These are essential metabolites that can be found through our saliva, blood, and urine. We even look at all the different varieties of estrogens which get broken down by the liver.
So, it�s fundamental that we look carefully at our hormone levels. Testing for hormonal imbalances in both men and women can tell us a lot about what�s causing our symptoms as well as what we can do to treat them. We can recommend a series of lifestyle modifications, including guidance and advice in nutrition and exercise. At least that�s what a good functional medicine doctor would do.
The scope of our information is limited to chiropractic and spinal health issues as well as functional medicine topics and discussions. To further discuss the subject matter, please feel free to ask Dr. Alex Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topic Discussion:�Acute Back Pain
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief. �
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download
* All the above XYMOGEN policies remain strictly in force.
El Paso, Tx. Chiropractor, Dr. Alex Jimenez presents the “Fasting Mimicking Diet�” (FMD�) by ProLon�. He introduces how the plan works, what it includes, and the benefits.
This 5-day meal program provides nutrients in precise quantities and combinations that nourish the body, but the body does not recognize�it as food and mimics a fast. This diet is the secret to fasting!
Research has shown certain types of diets that can mimic fasting, which enables the body to experience the health effects of a fast safely.
Fast Mimicking diet, what does it mean?
A Fasting Mimicking and Enhancing� Diet (FMED�) is a high nutrition, low protein, low carbohydrate meal plan, that benefits aging, poor health, inflammation and maintaining optimal health.
What does plan consist of?
The ProLon� plan is followed 5-days each month.
Suggested you follow a healthy diet for remaining 25 days.
Provides natural, healthy ingredients to nourish body while body believes it’s fasting.
Meal is low in carbohydrates & proteins
Contains healthy fatty acids
Plant-based soups
Bars
Crackers
Olives
Drinks
Supplements
How Diet Is Taken?
The diet should be taken for 5 consecutive days
Patient transitions one day then resumes normal diet gradually.
Specific combination of food provided for each day: Breakfast, Lunch, Dinner, and Snacks.
Missed meal can be made up any time same day.
Diet should be taken as recommended by healthcare professional.
After Completing the Diet?
6th-day diet ends, patient should avoid binge eating and resume normal diet gradually.
Should start with liquid foods:
Soups and fruit juices
Followed by light meals:
Rice, pasta and small portions of meat, fish
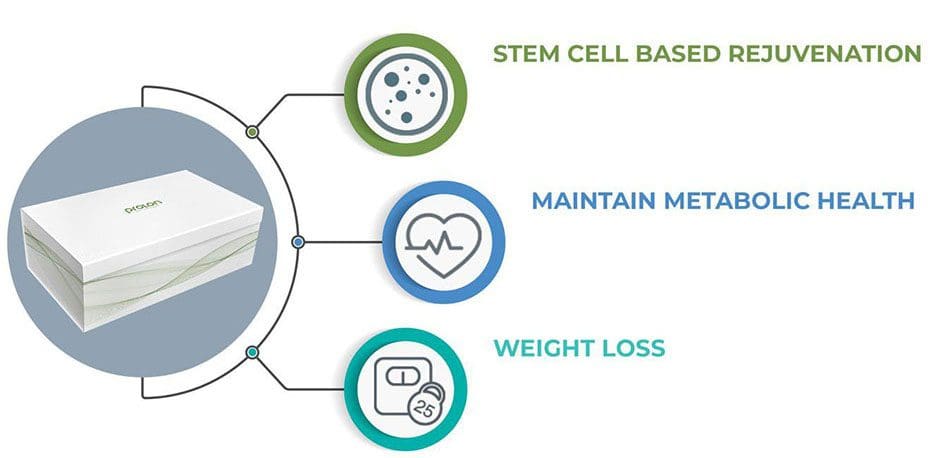
Body Performance Enhancement:
Allows body to trigger set of protection measures
Greater focus
Clarity
Energy
Leaner body
Decrease excess body fat
Preserve lean muscle mass
Fastest way to lose fat (belly fat)
Enhances cellular function
Promote stem cell-based renewal (cleans up aging & damaged cells)
Director of the Longevity Institute at the University of Southern California and The Program on Longevity and Cancer at IFOM in Milan designed the FMD.
He is considered the global leader in nutrition and aging.
His research team took on the journey to uncover an intervention that slows/reverses biological aging and delays the onset of age-related diseases.
Because it is risky nowadays to fast on water only, doctor Longo developed a natural plant-based meal program that imitates fasting while still feeding the body.
The ProLon Fasting Mimicking Formulation is only healthcare technology to be granted a patent for promoting tissue/organ regeneration, Longevity, and Healthspan by the USPTO.
You’ve seen body-weight scales at stores, online, in gyms and at the doctor’s office that utilize bioimpedance analysis. These scales can be expensive and wondered what is bioimpedance analysis and is it worth the price?
Bioelectrical impedance analysis may sound complicated, however, BIA devices use today’s technology. It measures the rate at which low-level electrical current is run through the body. Based on the rate that it travels, algorithms are used to measure fat-free mass, along with other data, such as height, gender, and weight measurements to determine your body fat percentage.
There are different types of devices, but each device requires two points of contact.
Handheld devices use two points, which are the hands (called hand-hand BIA).
Typical BIA scale uses the feet (called foot-foot BIA).
You place each foot on a pad and the current travels through your body between the feet.
There are also hand-to-foot BIA devices.
There are many brands that make different types of BIA scales (also called bioimpedance scales)
Newer models link with a smartphone app so you can track your progress.
Prices of a BIA scale depend on the sophistication of the equipment.
Some scales use multiple frequencies and more advanced algorithms, as well.
Some provide segmental fat analysis, which means, you can get body fat measurements for each arm, leg, and belly.
There are reports that segmental fat analysis (utilizing hand-foot BIA) are more accurate because hand-hand devices focus on measuring the upper body.
Foot-foot scales primarily measure the lower body.
These devices are considered safe for most people. Except:
Bioelectrical impedance analysis should not be used by anyone with an electronic medical implant, (e.g. heart pacemaker).
Not e used by pregnant women.
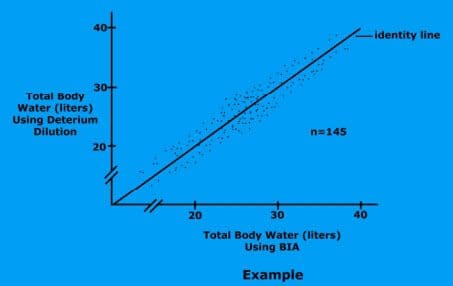
Studies have shown that bioelectrical impedance analysis is an accurate method for measuring body fat.
But these studies generally do not test the scales from the store.
Experts agree the accuracy of the measurement depends on the quality of the equipment.
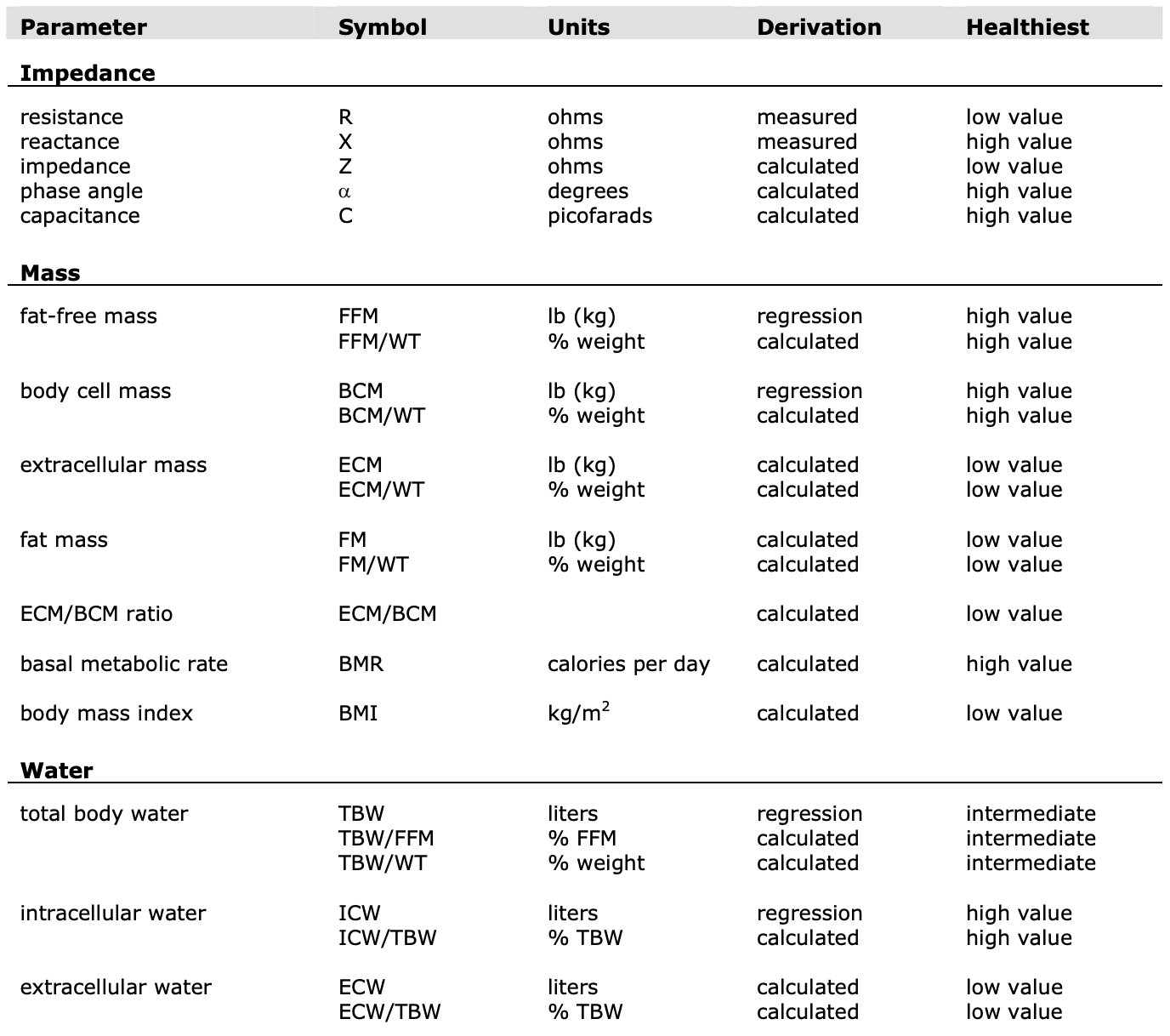
Parameters
(R) Resistance
Resistance and reactance are terms from physics, which are part of the field of materials and the effects on electricity. In reality, resistance and reactance are easy to understand.
Resistance is the ratio of electrical potential (voltage) to the current in a material. Put simply, a material with high resistance needs a high potential to generate a given amount of current in the material. A material with low resistance requires a low potential to produce the same amount of current in the material.
The easiest way to remember is:
Material with low resistance conducts well.
Material with high resistance conducts poorly.
When material conducts, it releases energy in the form of heat.
The resistance of a material is related to the material�s ability to dissipate energy.
Units of resistance are called ohms.
In the human body
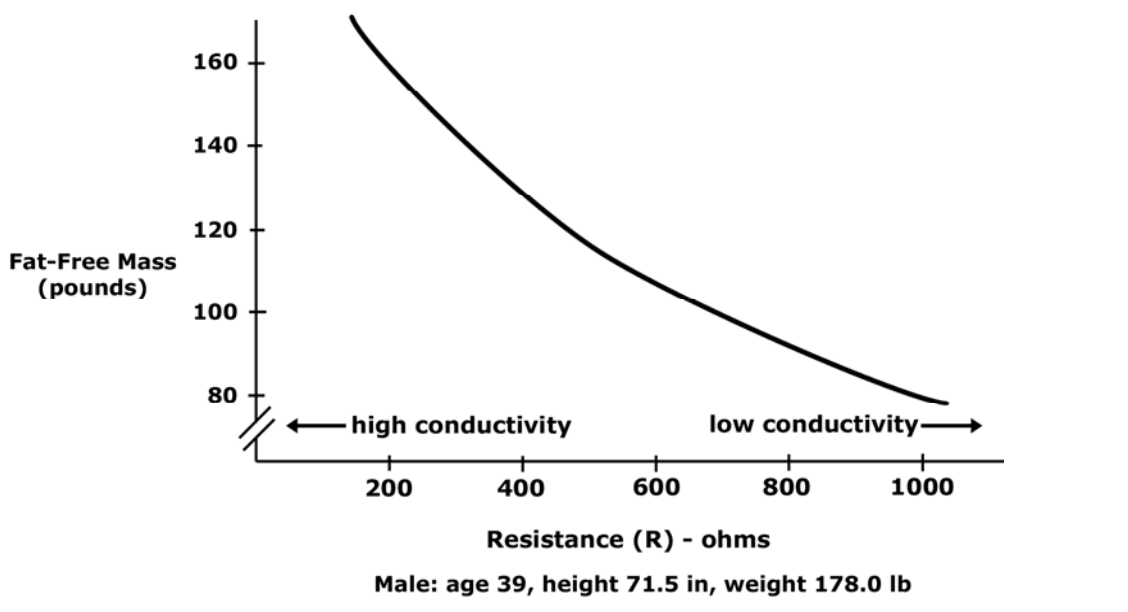
Low resistance is associated with large amounts of fat-free mass.
High resistance is associated with smaller amounts of fat-free mass.
The Case
Primary conductor in the human body is ionized water.
As the percentage of body weight that is water increases, the conductivity of the body increases.
Body water is contained solely in fat-free mass
Conductivity of body is proportional to amount of fat-free mass.
The potential required to generate the current is measured.
The ratio of potential and current along with a process called correlation and integration are used to determine the resistance and reactance. NOTE: This alternating current resistance is not the same resistance that can be measured by a standard store ohmmeter.
(X) Reactance
Reactance: the effect on an electrical current caused by a material’s ability to store energy.
A time delay between applied electrical potential and current.
Material that stores energy easily causes high reactance, and causes a large delay in the current.
Material that stores energy poorly has low reactance and causes a small delay in the current.
Example: Water poured onto top of a sponge will flow out of the bottom after a delay.
Large sponge will cause a large delay in the flow of water out the bottom
Small sponge would cause a small delay.
Current flows in material the same way.
The delay flow of current from storage is the reactance.
Units of reactance are ohms.
In the human body:
High reactance: large amounts of body cell mass (intracellular mass).
Low reactance: small amounts of body cell mass.
The Case
Cell membranes consist of a layer of nonconductive lipophilic material interposed between two layers of conductive molecules.
Behave like tiny capacitors and store energy.
Reactance in the body reflects the strength of capacitance.
Intact cellular membranes contained primarily within body cell mass
Reactance of the body is proportional to the amount of body cell mass
Reactance is Measured�by
Small current is applied through the body.
The potential required to generate the current is measured.
Ratio�of potential and current along with process correlation and integration used to determine the reactance.
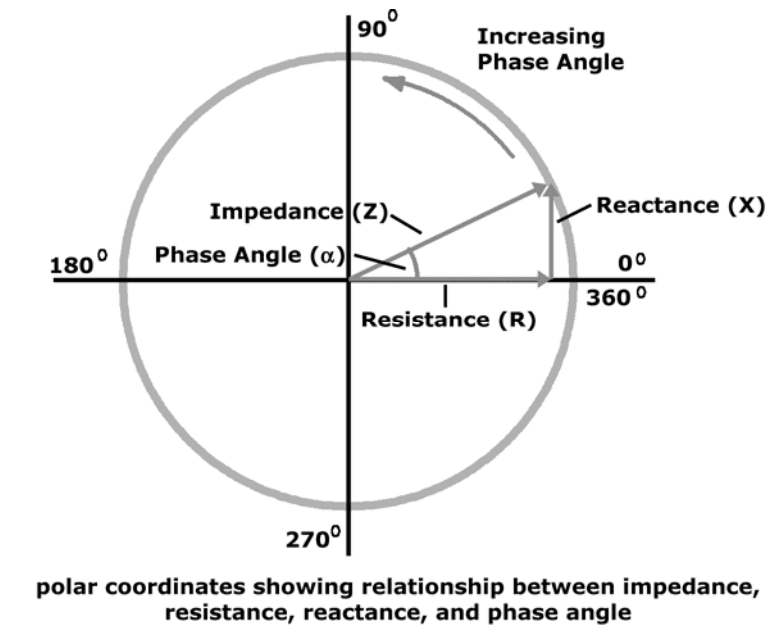
(Z) Impedance
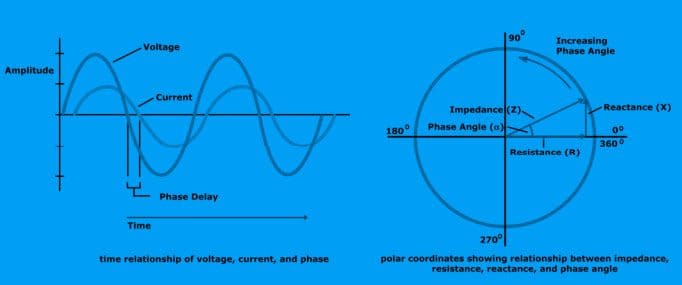
Total impedance (Z): The vector sum of effects of resistance and reactance on a current in the human body.
Technically, impedance is the ratio of potential (V) to current (I) and is used in bioimpedance analysis.
How R, X, & Z Relate
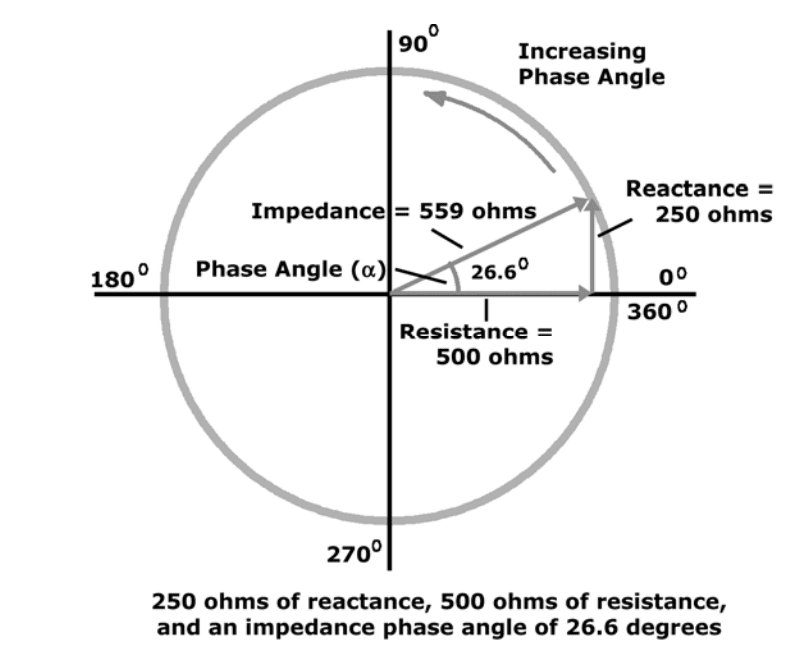
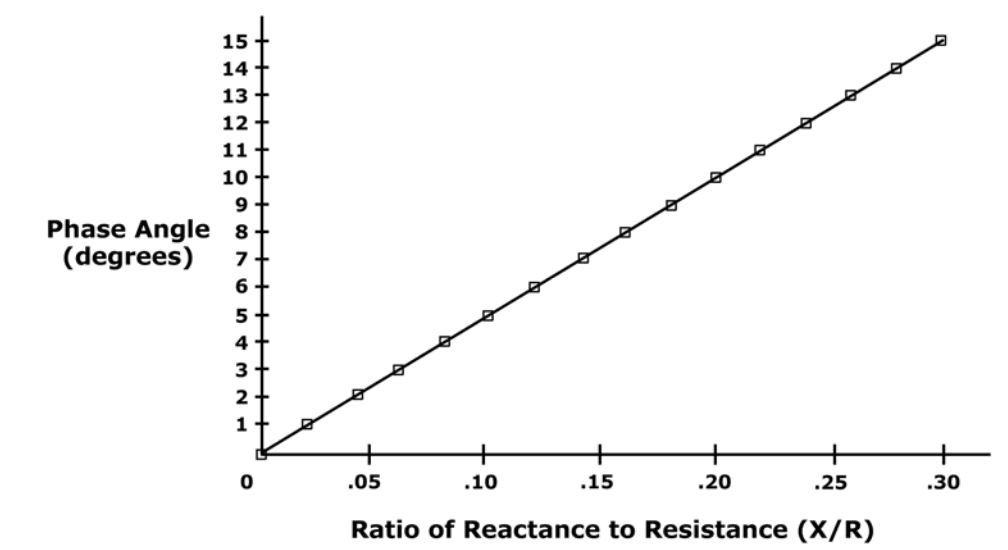
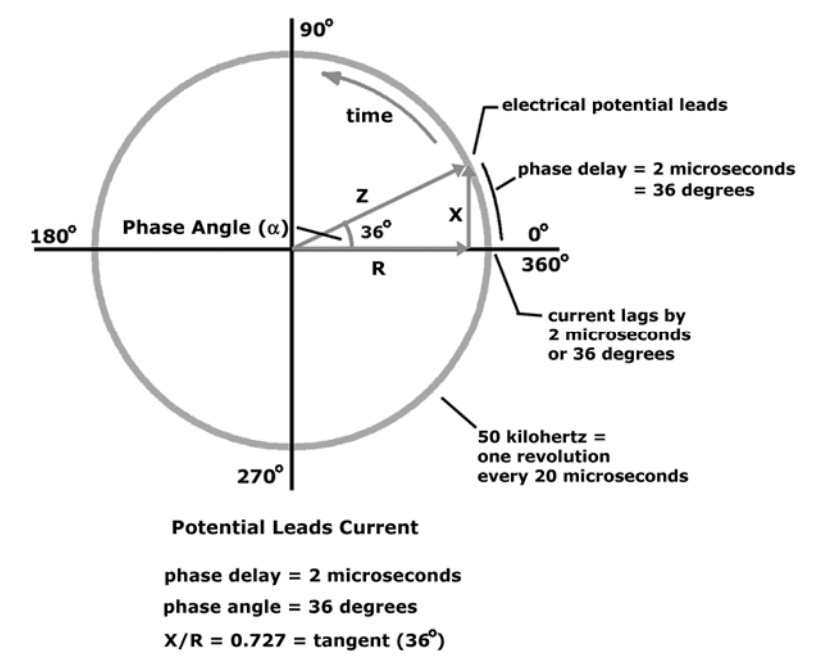
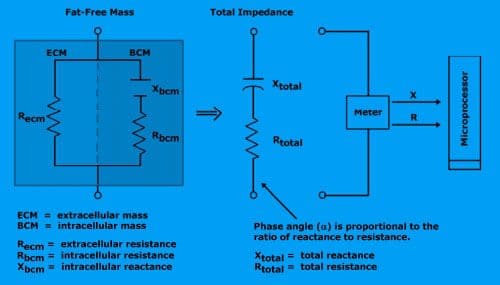
Mathematical relationship between Resistance (R), Reactance (X), and Impedance (Z) are: Z = sqrt (X2+R2) X = Z * sin (?) R = Z * cos (?) phase angle = arcsin (X/Z) phase angle = arctan (X/R)
Example:
(?) Phase Angle
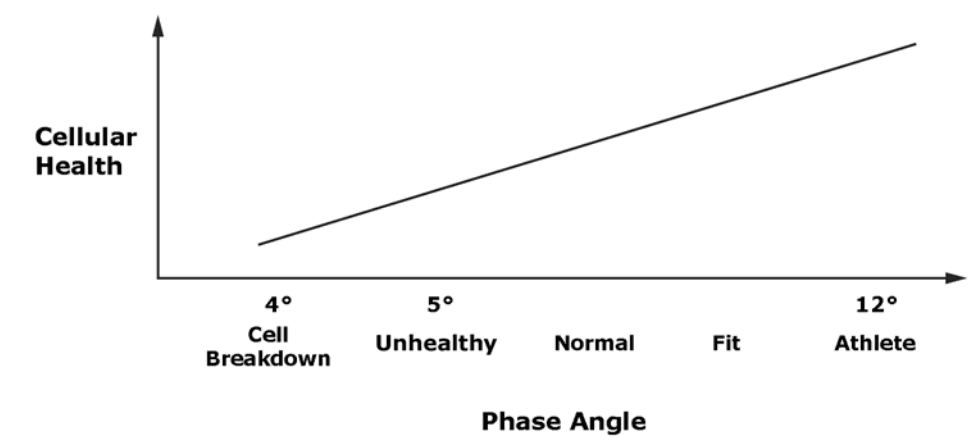
Phase angle is indicator of cellular health and integrity.
Research has shown the relationship between phase angle and cellular health is increasing and nearly linear.
Low phase angle: Consistent with inability of cells to store energy and
Indication of a breakdown in cellular membranes.
High phase angle: Consistent with large quantities of intact cell membranes and body cell mass.
Phase angle reflects the ratio of body cell mass to fat-free mass.
Phase angle is proportional to the ratio of reactance and resistance (proportional body cell mass to fat-free mass.)
Phase Angle Increase
Increase in body cell mass relative to fat-free mass.
Increase in fat-free mass relative to body weight.
Improved hydration of fat-free mass
Phase Angle: Useful For Comparing
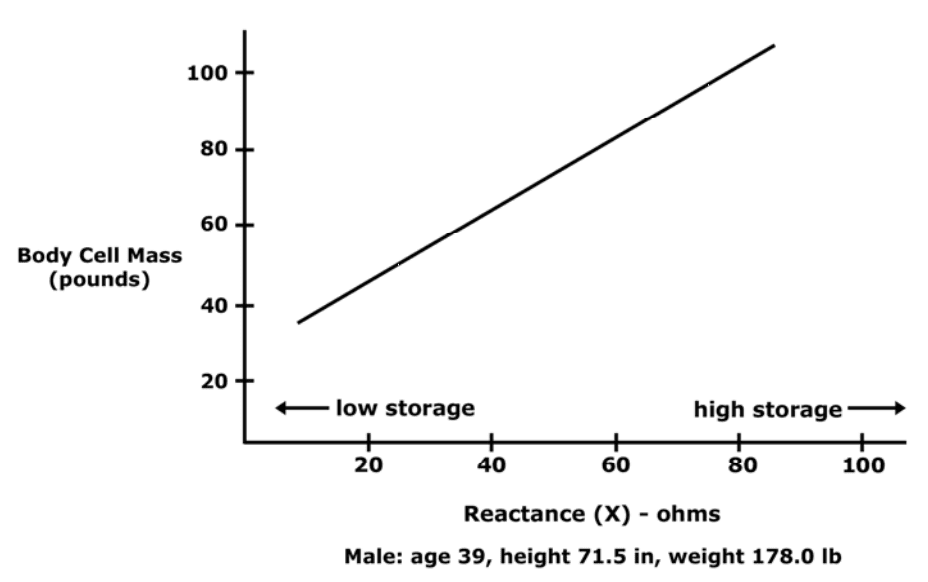
Reactance along with the patient’s weight indicates an absolute amount of body cell mass (BCM).
Reactance is best applied when comparing test results in a single patient at different times.
Two patients with exact reactance (X) can have different amounts of BCM, depends on patient’s weight.
Any patient with a higher phase angle will always have a higher proportion of BCM than patient with low phase angle.
Phase angle indicates number of intact cell membranes.
Phase angle is direct measurement of relative amounts of intact cellular membranes.
Phase Angle Works How?
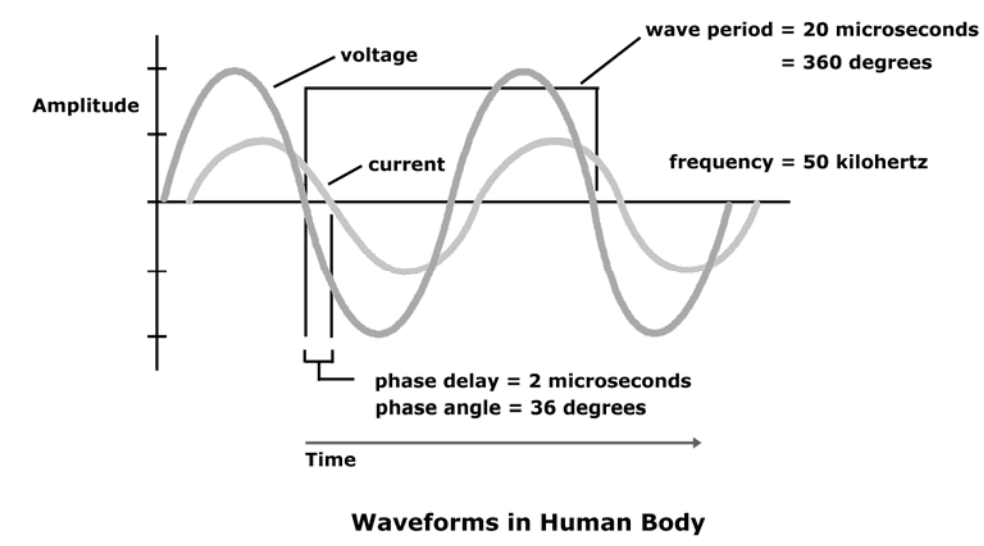
A bioimpedance analyzer applies 50-kilohertz alternating current to the body.
Phase angle is delay in time between the electric potential and current.
Oscilloscope connected to the body appears as delay between the voltage waveform and current waveform.
The period of each wave at 50 kilohertz is 20 microseconds. If, for example, the time delay is ten percent of the
period, then the time delay is 2 microseconds.
When expressed in units of time, it is said that the phase delay is 2 microseconds.
Time delay can be seen as percentage of entire wave period in degrees.
Complete wave period consists of 360 degrees. If time delay is one-tenth the total period of the wave, it is equivalent to 36 degrees.
When time delay is expressed this way (in degrees of total wave period),
This is the phase angle.
When electrical potential and current are depicted sweeping around a circle
Instead of moving over time
The relationship between reactance, resistance, and phase angle is easier to see.
Shown below
Range of phase angle in the human body is 1 to 20 degrees.
Phase angle is the arctangent of (X/R)
References:
Kyle UG, et al. Fat-Free and Fat Mass Percentiles in 5225 Healthy Subjects Aged 15 to 98 Years. Nutrition, 17:534-541, 2000.
Mattar J, et al. Application of total body bioimpedance to the critically ill patient. New Horizons 1995, Volume 4, No, 4: 493-503.
Ott M, et al. Bioelectrical impedance analysis as a predictor of survival in patients with human immunodeficiency virus infection. Journal of Acquired Immune Deficiency Syndrome and Human Retrovirology 1995: 9:20-25.
Functional Medicine Doctor Explains Men’s Hormones
Hello there, it�s Dr. Alex Jimenez again and welcome to part four of �Taking Control of your Healthcare�. Today, we�re going to discuss hormones. Hormones regulate most of the human body�s systems as they are recognized as the messenger molecules of the endocrine system. Hormone imbalances can cause subtle changes, however, their effects can tremendously impact an individual�s overall health and wellness. And what�s worse, most healthcare professionals don�t treat hormonal imbalances unless they�ve already been considered extreme.
The sex hormones, the thyroid hormones, and the adrenal hormones are the most important hormones we have to keep in balance. There�s a variety of other hormones, of course, but health issues associated with those are often more rare. Many doctors generally won�t test a person for hormone imbalances unless they�ve been trying to conceive a baby or they have sexual dysfunction or any other health issue of this type. And frequently, many doctors miss other problems by performing a screening test rather than a complete test.
What are Hormones?
Hormones are essential towards mental health, gut health, and reproductive health. In functional medicine, we even believe that hormones are vital towards maintaining a healthy immune system. Even if you�re not trying to conceive a baby or if you don�t have sexual dysfunction, it�s important for both men and women to known what their hormone levels look like.
Now, let�s discuss hormonal imbalances in sex hormones. First of all, How does a hormone imbalance manifest itself? Do you experience mood swings and fluctuations in your energy levels? Or if you�re a female, do you experience symptoms of PMS? Or perhaps you need coffee to wake up in the morning and wine to sleep at night? Has your sex drive, or libido, decreased? Do you have brain fog? Or is it difficult for you to focus on tasks?
If you�ve experienced any of these symptoms, you might have a hormonal imbalance in your sex hormones. Hormones are small molecules in charge of transmitting signals from one body system to another. But, if these molecules aren�t functioning accordingly, our energy levels and our mood can be tremendously affected. If you feel that something is off within your own body, talk to your doctor and seek proper testing. Don�t simply guess.
Let�s begin by discussing how you test your hormones. In functional medicine, we can test through saliva, blood, urine, and even stool. But, which is best for testing hormones? The truth is, knowing how and when to test your hormones is important because testing can depend on the marker that you want to look at. By way of instance, blood testing is one of the best and most affordable if you�re simply looking for a preliminary screening. A functional medicine doctor can also determine when a urine test will be more helpful than a blood test or a saliva test.
Taking Control of Men’s Hormones
So, now that you know how to test your hormones, we�ll discuss the different health issues caused by hormonal imbalances in the sex hormones of both men and women. First, let�s talk about men. As for the women, this is still an essential topic for you to learn about because the men in your lives are much less likely to schedule a doctor�s appointment for themselves.
Approximately 39 percent of men over the age of 45 have low levels of testosterone, or what�s known as low T. Elevated insulin levels associated with diabetes and obesity can lower your testosterone levels, which in turn, can increase estrogen levels. When testosterone goes down, your sex drive, or libido, and other functions can be tremendously affected. Excess consumption of sugar and starch can have different impacts for both men and women. Additionally, the utilization of certain drugs and/or medications, lack of exercise and physical activity, as well as inflammation, can also contribute to lower testosterone levels. While the majority of doctors won�t test testosterone levels in a man unless they have sexual dysfunction, I always want to make sure to test the sex hormones if a man has belly fat.
I would also like to discuss the serious side effects of a drug/medication which is commonly prescribed for sexual dysfunction. Statins can help lower cholesterol, however, did you know that your testosterone is made from cholesterol? That�s right. And when you start taking a drug/medication which was designed to decrease your cholesterol, you may also potentially be decreasing your sex hormones. It�s well-known that statins can decrease testosterone, leading to sexual dysfunction and even sometimes causing men to grow breasts, a condition known as gynecomastia. Side effects like these occur when we don�t treat the source of a health issue.
What you eat, including mainly sugar and starch, can be associated with your low testosterone levels and your abnormal cholesterol levels. Treatments like these where you take a drug for one thing but then end up taking another medication for the side effects of the fist medicine is unfortunately something that happens a lot in the medical field, and it can be a real nightmare.
Male testosterone levels are decreasing so much with each generation that normal reference ranges for testosterone levels in males are changing. But, that�s not something we want to happen. We don�t want these abnormal changes to become normal. Therefore, shouldn�t we be trying to find out why the overall health and wellness of our population is decreasing at such an alarming rate rather than lowering our normal standards of well-being?
Before we do that, however, we first have to understand why this problem is happening. From our increased exposure to toxins and our elevated levels of stress to our higher consumption of processed foods, these are only several of the more obvious reasons why our hormones are being affected.
In conventional medicine, the reference ranges for testosterone are between 264 and 916.
When you think about it, however, this is a tremendous range. Does a man with a testosterone level of 265 have the same sexual function as a man with a testosterone level of 916? Most definitely not. Yet these two people are classified under the same category. And with that in mind, what are the optimal reference ranges for testosterone? Men under the age of 30 should have a testosterone level of over 700, and men over the age of 30 should have a testosterone level of at least 500.
Evaluating men�s total testosterone levels is essential but we also need to evaluate their active hormone levels, or their free testosterone levels. Testosterone is carried around on what is known as a sex hormone binding globulin, which then releases it as the human body needs it. This carrier protein is found in the blood and when there�s too much of it, it becomes difficult for the human body to release testosterone when it�s needed.
In men, free testosterone levels should be of at least 10 but, they should optimally be closer to 15 or 20. Additionally, your doctor should check the sex hormone binding globulin or SHBG. As we discussed before, this carrier protein for testosterone and other hormones, can decrease your free testosterone levels. If you don�t check your SHBG, your total testosterone levels might be normal but your free testosterone levels might be too low.
By way of instance, SHBG is similar to a bus filled with many workers. In this case, the workers are testosterone. When we have too many buses, the majority of the workers will stay on the bus while only a few will be out doing their job. A man can have a total testosterone level of 700, however, if they have a free testosterone level of only 5, they�re bound to still feel like they have a total testosterone level of 300.
In summary, we want to make sure that our total testosterone level is over 500 or better, over 600 and we also want to make sure that our free testosterone level is between 15 and 20.
Another hormone you might want to make sure you get tested for is known as dehydroepiandrosterone, or DHEA. DHEA is a precursor hormone for testosterone. It�s also an adrenal hormone, however, we will discuss this later in another article. If an individual�s DHEA is too low, it can indicate that the adrenal glands, which are in charge of the human body�s stress response, may not be functioning appropriately. DHEA levels should be between two to 400.
DHEA, or dehydroepiandrosterone, can be supplemented directly. Many doctors and functional medicine practitioners may also implement stress-relieving methods and techniques into your life, such as mindfulness meditation and yoga, to treat the source of the health issue. Herbal supplements can also help regulate DHEA as well as testosterone levels.
Now, we will discuss a fundamental hormone that is generally only considered to be important in female health, however, this hormone also plays an essential role in men; estrogen. Estrogen helps maintain a healthy sex function as it promotes your libido. It is also protective for the brain. In men�s health, estrogen is often demonized because it can cause health issues like breast development if estrogen levels are too high in men. But, normal estrogen levels are fundamental towards maintaining hormone balance as well as mental health.
Estrogen can increase in men with diabetes and obesity. Elevated insulin exchanges more testosterone into estrogen which may cause additional symptoms like fatigue. This can also create more problems alongside sexual dysfunction, including hair loss. So, if you have less than optimal testosterone levels, you should seek help from a doctor or functional medicine practitioner to find out why. And looking at your estrogen levels is a great place to start.
There are two types of estrogen tests which can help demonstrate your estradiol and estrone levels. These are important markers to look at because one can be elevated while the other can be in the normal range. The brain is the other place you can look at to determine your estrogen levels. Your brain produces hormones which can stimulate sex hormones, such as the follicle stimulating hormone, or FSH, and the luteinizing hormone, or LH, which helps produce testosterone and sperm within the testicles. If your LH is low, then your low testosterone levels may be caused by a brain health issue. However, it�s much more likely that sugar and starch consumption may be causing hormonal imbalances in your sex hormones.
Approximately 70 percent of the testosterone deficiencies in the United States are associated with insulin resistance due to diabetes and obesity. So, if your diet is filled with sugar and starch or if you have belly fat, you may already have decreased testosterone levels.
Hormones are secreted directly into the blood stream in order to control a variety of bodily functions. These can affect growth and development, mood, sexual function, reproduction, and metabolism. As a fundamental part of the endocrine system, hormone imbalances can have a tremendous effect on our overall health and wellness. Men’s hormones, by way of instance, can significantly impact a man’s quality of life. Research studies have demonstrated that decreased testosterone in men can cause a variety of health issues. Evaluating sex hormones in both men and women is essential towards overall health and wellness. Dr. Alex Jimenez D.C., C.C.S.T. Insight
Understanding Men’s Hormones
While we keep discussing the importance of lab tests, we also need to keep asking ourselves why these health issues are happening in the first place. The answers trace back to the basics of health and wellness. What are you eating? Do you participate in exercise or physical activities? Do you sleep properly? Do you have stress? What are your nutrient levels? Of course, several of these answers require more evaluations. That�s why I highly recommend you find a functional medicine doctor who can help get you on the right path towards optimal health and wellness. Most of the time, basic lifestyle modifications including diet, exercise, stress management, sleep and supplements can help. Other times you may need hormone replacement therapy.
The scope of our information is limited to chiropractic and spinal health issues as well as functional medicine topics and discussions. To further discuss the subject matter, please feel free to ask Dr. Alex Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topic Discussion:�Acute Back Pain
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief.
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download
* All the above XYMOGEN policies remain strictly in force.
Bioimpedance Analysis (BIA) is defined as the analysis of resistance and reactance in the human body.
Energy dissipation is called resistance (R).
Energy storage is called reactance (X).
Units of resistance and reactance are called ohms.
Resistance and reactance taken together create impedance (Z).
Fat-free mass in the human body is proportional to its resistance.
Body cell mass is proportional to its reactance.
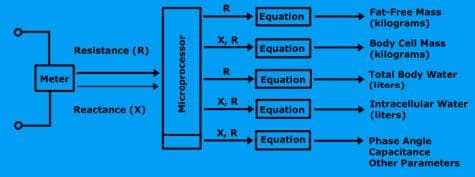
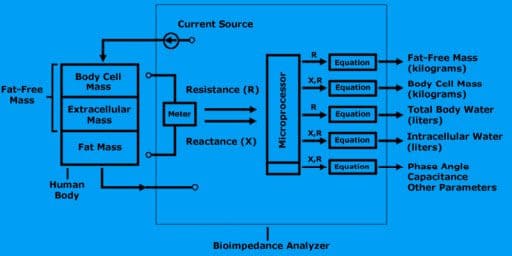
A bioimpedance analyzer is a device that contains a highly precise frequency meter, a processor, and a printer. The meter is called an impedance meter. It measures resistance and reactance.
The microprocessor computes the size of the fat-free mass and body cell mass along with all subsequent calculations.
A bioimpedance analyzer measures resistance and reactance, computes fat-free mass, body cell mass, total body water, and intracellular water.
To measure resistance and reactance, the bioimpedance analyzer generates a small electrical current to the body.
Measurements
Bioimpedance analysis is an assessment of change in electrical tissue conductivity, which indicates altered body composition.
The following electrical characteristics are measured and used to evaluate body composition.
Impedance (Z)
Impedance is a measurement of a medium to conduct current. It is a ratio of voltage injected current in a conductive medium and has two components: resistance and reactance.
Resistance (R)
Resistance is the component of impedance related to the dissipation of energy.
Reactance (X)
Reactance is the component of impedance related to the storage of energy.
Phase angle is a time delay between stimulating current and the voltage generated by an alternating current. The phase angle is expressed in degrees.
Electrical characteristics and their relation to each other?
The time relationship of voltage, current, and phase is shown below.
The Circuit
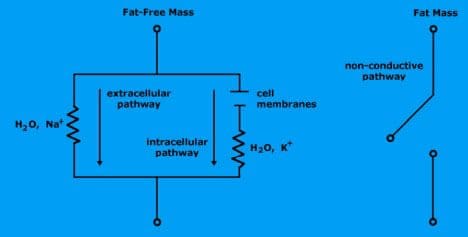
Water is the body’s conductor and determines the resistance.
Electrolytic fluid in fat-free mass consists of water and charged ions that conduct electrical current.
Extracellular fluid (water and ionized sodium Na+) provides a low-resistance pathway.
Intracellular fluid (water and ionized potassium K+) provides a low-resistance pathway.
Fat mass is non-conductive because there is no water.
Cell membranes within intracellular mass determine reactance.
The cell membranes consist of a layer of nonconductive, lipophilic material interposed between two layers of conductive molecules. They are tiny capacitors that store electrical charge.
Extracellular pathway (which is sodium ionized water) is represented by single resistance.
Intracellular pathway (potassium ionized water and cell membranes) is represented by resistance and reactance.
Total resistance is proportional to the amount of fat-free mass.
Reactance is proportional to body cell mass (intracellular mass).
The meter reports total resistance and total reactance to the microprocessor.
Equation for Regression
Bioimpedance analyzers use algorithms to compute amounts of fat-free mass, body cell mass, body water, and intracellular water.
What is the regression equation?
This equation is a mathematical formula that predicts the relationship between two or more variables based on the analysis of data.
For example, total body water�regression�equation
(TBW) = form:
TBW = a * Ht2 / R + b * Weight + c * Age + d
The terms are Ht2 / R, Weight, and Age.
The weighting constants are a, b, c, and d.
Resistance, height, weight, gender, and age are supplied for each subject. A computer then generates an algorithm which best describes the relationship between TBW, resistance, height, weight, gender, and age.
This is called Regression Analysis.
How do you use regression equations?
Once the bioimpedance analyzer measures the resistance and reactance, the results are sent to the microprocessor to perform the calculations. All the equations, (FFM, BCM, TBW, and ICW) are encoded into the analyzer’s software.
Diagram
Fat-Free Mass
Fat-free mass is derived by applying a small alternating current to the body.
The resistance of the body is measured using the measured resistance (R), the analyzer estimates the fat-free mass.
Body Cell Mass
Body cell mass is derived by measuring both the resistance (R) and reactance (X) of the body. With this measurement, the analyzer estimates body cell mass.
Total Body Water
Using the measured resistance, the analyzer estimates total body water.
Intracellular Water
Using the measured resistance and reactance, the analyzer estimates intracellular water.
El Doctor de Medicina Funcional Explica la Nutrici�n
Cada reacci�n qu�mica que ocurre en el cuerpo humano requiere de enzimas y cada uno de estos procesos necesita una coenzima. Pero �qu� son las coenzimas? Son vitaminas y minerales. Aproximadamente 37 billones, billones de reacciones qu�micas ocurren en el cuerpo humano cada segundo.
Es por eso que una nutrici�n adecuada y una dieta balanceada rica en alimentos integrales con vitaminas y minerales es fundamental para la salud y el bienestar general. La mayor�a de las personas en los Estados Unidos son deficientes en vitaminas y / o minerales. Pero, �c�mo sabes si eres parte del 90 por ciento de las personas con suficientes deficiencias para desarrollar una enfermedad? Discutiremos las pruebas que puede realizar para averiguar si tiene deficiencia de vitaminas y / o minerales y qu� puede hacer al respecto.
�Que es la Nutrici�n?
Hola, bienvenidos a la tercera parte de “C�mo Tomar el Control de su Salud”. Hoy, discutiremos uno de los temas mas divertidos de la medicina funcional: la nutrici�n. Desafortunadamente, la nutrici�n es una de las conversaciones m�s importantes que muchos doctores no est�n dispuestos a tener con sus pacientes. El doctor promedio aprende sobre las enfermedades y la desnutrici�n en lugar de aprender c�mo usar la nutrici�n como tratamiento o incluso c�mo usar terapias nutricionales para lograr salud y bienestar �ptimos.
Personalmente creo que la comida puede ser utilizada como una forma de medicina. Que deber�a ser la base de la pr�ctica m�dica, no una idea tard�a en la medicina. No hay mejor tratamiento que la nutrici�n adecuada. Aproximadamente el 90 por ciento de las personas en los Estados Unidos no obtienen los nutrientes esenciales que necesitan para las funciones corporales saludables. Y m�s que eso probablemente no est� obteniendo suficientes nutrientes para prevenir enfermedades asociadas con deficiencias nutricionales. Sin embargo, �qu� se necesita para lograr un bienestar �ptimo? M�s del 98 por ciento de los estadounidenses son deficientes en omega-3, 80 por ciento en vitamina D, 50 por ciento en magnesio y 10 por ciento en vitamina C. Las deficiencias de nutrientes tambi�n pueden seguir causando problemas de salud durante a�os.
Las enfermedades agudas, como el raquitismo, el escorbuto, el beriberi o la anemia por deficiencia de hierro, suelen ser los problemas de salud m�s comentados asociados con las deficiencias de nutrientes. Sin embargo, tambi�n se conocen como enfermedades de deficiencia de latencia prolongada. Entonces, �cu�nta vitamina D necesitamos para no tener raquitismo? No mucho, solo 30 unidades realmente. �Y cu�nto necesitamos para no tener osteoporosis? Tal vez unas 3,000 a 4,000 unidades por d�a. Ahora, �cu�nto folato necesitamos para no tener anemia? Tambi�n no mucho. Pero, �cu�nto necesitamos para prevenir las enfermedades card�acas, el c�ncer y la demencia? Definitivamente se necesitan muchas m�s unidades por d�a.
Cada reacci�n qu�mica que ocurre en el cuerpo humano requiere enzimas y cada uno de estos procesos necesita una coenzima. Pero �qu� son las coenzimas? Son vitaminas y minerales. Aproximadamente 37 billones, billones de reacciones qu�micas ocurren en el cuerpo humano cada segundo.
Es por eso que una nutrici�n adecuada y una dieta balanceada rica en alimentos integrales con vitaminas y minerales es fundamental para la salud y el bienestar general. La mayor�a de las personas en los Estados Unidos son deficientes en vitaminas y / o minerales. Pero, �c�mo sabes si eres parte del 90 por ciento de las personas con suficientes deficiencias para desarrollar una enfermedad? S�lo hay varios nutrientes que generalmente se analizan. Y para la mayor�a de estos, los doctores no son conscientes de cu�les deber�an ser los valores �ptimos, lo que puede dificultar mucho la correcci�n de la deficiencia de nutrientes.
Tomando el Control de su Nutrici�n
Uno de los nutrientes m�s fundamentales que se necesita medir es la vitamina D. Aunque se le conoce como una vitamina, en realidad es m�s como una hormona y se produce por parte del colesterol. Esta es otra raz�n por la cual el colesterol es esencial. Aproximadamente el 80 por ciento de la poblaci�n tiene una deficiencia de vitamina D. A menos que est� al sol 20 minutos todos los d�as entre las 10:00 am y las 2:00 pm, es posible que deba tomar suplementos de vitamina D. Para complementar adecuadamente, necesitamos saber desde qu� nivel est� comenzando desde un principio. A modo de ejemplo, los niveles �ptimos de vitamina D deben de ser entre 50 y 80 nanogramos por mililitro de sangre. La cantidad recomendada de vitamina D que podemos complementar es de aproximadamente 2,000 a 4,000 unidades.
Si tiene niveles m�s bajos de vitamina D o si tiene problemas gen�ticos, es posible que necesite un suplemento con hasta 10,000 unidades de vitamina D. Por eso es fundamental trabajar con un doctor o especialista de medicina funcional que pueda medir y evaluar sus niveles de nutrients, as� como ayudarles a optimizarlos. La mayor�a de los suplementos contienen aproximadamente 400 unidades, que es 10 veces menos que la cantidad que la mayor�a de nosotros necesitamos. Los niveles comunes son generalmente un poco m�s de 20. Esto es demasiado bajo. En un estudio de investigaci�n, las mujeres con niveles de vitamina D entre 45 y 60 experimentaron una reducci�n en los partos prematuros hasta de un 60 por ciento. La vitamina D tambi�n es esencial para ayudar a desarrollar huesos y m�sculos fuertes, mejorar la funci�n del sistema inmunol�gico, prevenir el c�ncer asi como para ayudarlo a vivir m�s tiempo. Es incre�ble.
Otra medida o prueba realizada por la mayor�a de los doctores, pero que no siempre se interpreta correctamente, se conoce como MCV o volumen corpuscular medio. La medida de MCV eval�a el tama�o de sus gl�bulos rojos en una prueba llamada CBC, o hemograma completo, que es uno de los paneles de sangre m�s comunes solicitados por profesionales de la salud. Por lo tanto, si usted es deficiente en nutrientes, sus c�lulas pueden hacerse m�s peque�as o m�s grandes. A modo de ejemplo, si sus c�lulas son demasiado grandes, podr�an ser signos de una deficiencia de folato o vitamina B12.
Las vitaminas B son esenciales en numerosas reacciones qu�micas en el cuerpo humano. Nos ayudan a producir energ�a y nos ayudan a regular la expresi�n gen�tica para crear prote�nas que aseguran nuestra salud y bienestar en general. Si nuestras vitaminas B estan demasiado bajas, eventualmente podr�amos desarrollar una deficiencia de hierro, anemia o incluso podr�a causar un trastorno gen�tico.
Los niveles �ptimos de vitaminas B deben estar entre 80 y 90. Los suplementos vitam�nicos del complejo B pueden ayudar a optimizar f�cilmente los niveles de vitaminas B. Pero, �por qu� alguien ser�a deficiente en vitaminas B? �Acaso su dieta no les proporciona suficientes nutrientes? �Son veganos? �Est�n tomando medicamentos que evitan la absorci�n de la vitamina B12? Adem�s, las vitaminas B se agotan en momentos de gran estr�s que, como quiropr�ctico, puedo decir que le sucede con frecuencia a la mayor�a de la poblaci�n en los Estados Unidos.
El MCV no es la �nica medida o prueba que eval�a los niveles de vitaminas B de un paciente. La homociste�na es un marcador alternativo que analizaremos en futuros art�culos que demuestran los niveles de B6, folato y B12. Sin embargo, tanto el MCV como la medida o prueba de homociste�na solo demuestran que uno o m�s de estos nutrientes pueden ser deficientes. No necesariamente nos dice cu�l. Por lo tanto, se pueden requerir algunas evaluaciones adicionales de seguimiento.
La medida o prueba del MMA, o �cido metilmal�nico, tambi�n muestra los niveles de vitamina B12. La vitamina B12 es esencial para muchos procesos en el cuerpo humano, incluida la producci�n de energ�a, la expresi�n de genes, la metilaci�n, la funci�n nerviosa y el estado de �nimo, entre muchos otros procesos. Los veganos tienen una mayor probabilidad de desarrollar una deficiencia de B12 porque este solo se encuentra en los productos animales. El folato es otra vitamina B fundamental. Se puede determinar directamente en la sangre, pero la homociste�na es un marcador m�s preciso para los niveles de folato.
En esta secci�n, tambi�n hablaremos sobre gen�tica porque existe una medida o prueba que puede demostrar mucho m�s con respecto al estado de sus vitaminas B y su capacidad para utilizarlas. Nuestros genes son capaces de producir prote�nas. Tenemos aproximadamente 20,000 genes que est�n dise�ados para crear prote�nas. Y un tercio de todas las prote�nas que producen son para nuestras enzimas. Las enzimas convierten los nutrientes en otros nutrientes. Estas enzimas tambi�n dependen en gran cantidad de ciertos nutrientes espec�ficos. Uno de los genes m�s fundamentales que pueden verse afectados es conocido como MTHFR o metilentetrahidrofolato reductasa. Pero puedes llamarlo MTHFR.
El MTHFR es esencial porque ayuda a regular la metilaci�n, la homociste�na y el folato, que son vitales para nuestra salud y bienestar general. Cuando se tienen niveles elevados de homociste�na, se debe verificar su estado de metilaci�n buscando el gen MTHFR a trav�s de un simple an�lisis de sangre.
La metilaci�n es un proceso bioqu�mico clave que es fundamental para la funci�n adecuada de la mayor�a de los sistemas del cuerpo humano. Se dispara miles de millones de veces cada segundo. Y ayuda a controlar la homociste�na, una sustancia que puede da�ar los vasos sangu�neos y se ha asociado con la demencia, asi como enfermedades card�acas y c�ncer, entre otros problemas de salud. La metilaci�n tambi�n ayuda a reparar su ADN de manera regular, ya que ayuda a reciclar las mol�culas necesarias para la desintoxicaci�n o para eliminar las toxinas. Tambi�n ayuda a controlar su estado de �nimo y ayuda a controlar la inflamaci�n. La metilaci�n es vital.
Pero, para asegurarse de que su metilaci�n est� activa, el cuerpo humano necesita niveles �ptimos de vitaminas B. Sin suficientes vitaminas B, el proceso de metilaci�n se puede descomponer y los efectos pueden ser destructivos. Aqu� es donde comenzamos a ver un aumento en los defectos cong�nitos, como la espina b�fida, el s�ndrome de down y m�s abortos involuntarios.
MTHFR es frecuentemente anormal en aproximadamente el 35 por ciento de la poblaci�n. La descomposici�n de la metilaci�n tambi�n puede aumentar el riesgo de desarrollar problemas de salud como la osteoporosis y la diabetes, la displasia cervical o el c�ncer, incluido el c�ncer de colon y el c�ncer de pulm�n, e incluso la depresi�n, la disfunci�n cognitiva pedi�trica, as� como los trastornos del humor y del comportamiento, la demencia y los problemas cerebrovasculares. La metilaci�n es verdaderamente un proceso bioqu�mico clave.
Cuando hablamos de la gen�tica, debemos entender que nuestro entorno puede alterar nuestros genes. Entonces, �qu� pasa si tienes una variaci�n MTHFR en tus genes? En primer lugar, no todas las mutaciones causan problemas de salud. Una mutaci�n, por modo de ejemplo, conocida como C677T, es una versi�n del gen que es m�s significativa que otra versi�n del gen, conocida como A1298C. Ahora no hay que preocuparse por estas variaciones gen�ticas. Sirven como ejemplos para demostrarle la calidad de estas mutaciones y c�mo funcionan. Las personas con estas variaciones del gen, por ejemplo, solo pueden necesitar m�s folato o pueden necesitar un tipo particular de folato conocido como metilfolato. Aqu� es donde un practicante de medicina funcional puede ayudarle a sus pacientes.
Una prueba gen�tica puede hacerle saber si tiene una de estas variaciones gen�ticas. Pero, no te estreses. Hay mucho que puede hacer para optimizar su salud y bienestar en general. Muchos pacientes han visitado mi consultorio despu�s de descubrir que tienen estas variaciones en sus genes. Y aprenden r�pidamente que tienen la opci�n de tomar el control de su bienestar. Sin embargo, lo que controlas no son tus genes, controlas tu expresi�n gen�tica.
Si alteras tus h�bitos alimenticios, alteras tus nutrientes. Si altera su entorno, altera qu� genes se activan y cu�les se vuelven inactivos. Y con estas mutaciones, puedes hacer casi lo mismo simplemente siguiendo una nutrici�n adecuada. Cuando encuentre un m�dico o practicante de medicina funcional que est� dispuesto a trabajar con usted, le dir�n qu� modificaciones de estilo de vida debe seguir para prevenir problemas de salud.
Por lo tanto, acabamos de discutir las vitaminas B. A continuaci�n, discutiremos otro nutriente fundamental en el cuerpo humano: el magnesio. El magnesio es un mineral s�per esencial. Aproximadamente el 48 por ciento de las personas en los Estados Unidos consumen menos de la cantidad requerida de magnesio en los alimentos. El magnesio es necesario en m�s de 300 reacciones qu�micas en el cuerpo humano. Tambi�n es fundamental en la producci�n de ATP, o la energ�a que el cuerpo humano utiliza como combustible.
Una prueba o medida del nivel de magnesio en la sangre puede ayudar a determinar si usted tiene una deficiencia. El magnesio tambi�n puede ayudar a reducir la ansiedad, calmar el sistema nervioso y mejorar el sue�o. Tambi�n es un nutriente esencial en el manejo de los niveles de az�car en la sangre. Si un profesional de la salud le ha dicho que tiene un nivel promedio de az�car en la sangre de m�s de cinco y medio, algo conocido como A1c, entonces el magnesio puede ayudar.
Adem�s, es muy f�cil saber si tiene una deficiencia de magnesio al observar su dieta y s�ntomas actuales. �Come suficientes alimentos ricos en magnesio como verduras de hojas verdes, frijoles, nueces y semillas? �O come muchos alimentos procesados? Quiz�s tambi�n tenga s�ntomas como ansiedad, insomnio, estre�imiento, contracciones musculares, calambres musculares, s�ndrome premenstrual y palpitaciones. Si tiene uno o m�s de los s�ntomas que acabo de mencionar, es posible que tenga una deficiencia de magnesio.
A continuaci�n, hablaremos sobre el zinc, el mineral que estimula la inmunidad y la testosterona en el cuerpo humano. Este importante nutriente se encarga de mantener el volumen de su cabello, as� como de reparar su tracto gastrointestinal. Tambi�n es responsable de asegurarse de que su tiroides funcione correctamente. El zinc puede medirse o analizarse f�cilmente en la sangre y, desafortunadamente, es otro nutriente de el que tenemos una gran deficiencia en los Estados Unidos. Adem�s, tambi�n se pueden observar sus niveles de fosfatasa alcalina, que pueden calcularse a trav�s de una evaluaci�n de la funci�n hep�tica en un panel de sangre regular. Los niveles altos de fosfatasa alcalina pueden indicar la presencia de c�ncer o problemas �seos, entre otros problemas de salud; sin embargo, los niveles bajos de fosfatasa alcalina pueden indicar una deficiencia de zinc, ya que es una enzima dependiente de zinc.
Finalmente, el �ltimo nutriente fundamental de el que vamos a discutir es el hierro. El hierro es frecuentemente deficiente en veganos y vegetarianos, o en mujeres en general debido a la menstruaci�n. El hierro es necesario para transportar ox�geno a trav�s del cuerpo humano y es esencial para la salud y el bienestar del cerebro. El hierro tambi�n es importante para el cabello y las u�as, el sue�o y muchas otras cosas.
La ferritina es un tipo de hierro almacenado y es este nutriente el que ayuda a ver tus niveles de hierro. Los niveles �ptimos de ferritina deben estar entre 50 y 150 en las mujeres y entre 100 y 300 en los hombres. Muchas veces he visto a mujeres visitar mi oficina con niveles de ferritina de menos de 50, o peor, de un solo d�gito. Esto se debe a que las mujeres premenop�usicas pierden sangre cada mes debido a sus ciclos menstruales y se les hace mucho m�s dif�cil mantener los niveles adecuados de ferritina. Muchas mujeres tambi�n comen mucho menos de lo que se supone que deben comer todos los d�as. Los altos niveles de ferritina, por otro lado, podr�an ser un signo de inflamaci�n, generalmente causada por la resistencia de la insulina al az�car, o podr�a ser un signo de hemocromatosis o enfermedad por almacenamiento de hierro, un trastorno gen�tico que puede ser muy peligroso.
Tener niveles reducidos de ferritina tambi�n puede hacer que se sienta cansado, y puede causar p�rdida de cabello, al igual que puede causar insomnio. Entonces, incluso si su recuento sangu�neo es normal, si sus niveles de ferritina son bajos o sus niveles de hierro son bajos, tambi�n puede causar estos s�ntomas. Por eso, si experimenta s�ntomas de fatiga, es esencial medir o probar sus niveles de ferritina. Y se puede complementar f�cilmente.
Aparte de la ferritina, un MCV bajo tambi�n puede determinar si usted tiene una deficiencia de hierro. Las deficiencias de hierro pueden hacer que los gl�bulos rojos se vuelvan muy peque�os y eso se puede demostrar en los niveles bajos de MCV, que eval�an el tama�o de sus gl�bulos rojos. Adem�s, la saturaci�n de transferencia, el hierro s�rico, la TIBC o la capacidad de uni�n al hierro total, y la hemoglobina, pueden brindarnos un an�lisis m�s profundo del estado de su hierro para distinguir las diferentes causas de la anemia. Estos se incluyen en un panel de sangre de hierro regular en una prueba de laboratorio.
Hemos discutido varios nutrientes que pueden ser solicitados por la mayor�a de los profesionales de la salud con acceso a pruebas de laboratorio convencionales. Adem�s, hay otra prueba que nos puede dar m�s informaci�n sobre qu� tipo de nutrientes necesitamos en funci�n de nuestros genes. Se llama la prueba de salud de ADN y es proporcionada por una compa��a llamada DNAlife. Esta prueba eval�a una variedad de marcadores gen�ticos asociados con la desintoxicaci�n, el metabolismo de los l�pidos y la inflamaci�n, incluido el gen MTHFR y otros marcadores de vitamina B. Ahora, DNA Health demuestra los diferentes genes que evaluamos. Y la mayor�a de estos son genes comunes, son aquellos sobre los que podemos hacer algo. Analizamos los genes que podemos cambiar seg�n su nutrici�n y otros factores del estilo de vida.
El gen MTHFR nos muestra otros marcadores de vitamina B, genes que controlan B6, folato y B12, adem�s de demostrar c�mo funcionan y si usted tiene deficiencias de nutrientes. Luego, nos indica qu� nutrientes necesitar� complementar y cu�nto le daremos. Es tremendamente �til.
Hubo un individuo que ten�a dos variables del gen MTHFR. Esta mujer tuvo un aborto involuntario tras otro aborto involuntario tras otro aborto involuntario. Visit� a su m�dico para una evaluaci�n y result� que ten�a una mutaci�n reguladora del folato. Entonces, su m�dico comenz� a darle la cantidad adecuada de folato que necesitaba y ella comenz� a tener beb�s sanos. A veces, la nutrici�n puede ser asi de poderosa para mejorar la salud y el bienestar general del paciente.
La prueba de salud del ADN puede ayudar a personalizar su enfoque al optimizar su bienestar basado en su gen�tica. Lo que medimos utilizando la prueba de salud del ADN proporciona informaci�n bien establecida acerca de sus genes y lo que puede hacer al respecto.
Una prueba de micronutrientes conocida como el perfil de nutrici�n optimizado individualizado o el panel ION, son opciones de prueba alternativas que tambi�n pueden proporcionar informaci�n sobre su estado nutricional actual. Esta prueba es de Genova. Esta es una prueba s�lida que mide todas las vitaminas y minerales esenciales, �cidos grasos, �cidos org�nicos y antioxidantes que tiene actualmente. Esta prueba busca desequilibrios, insuficiencias o deficiencias, en lugar de buscar una enfermedad espec�fica. Busca cosas que la mayor�a de los m�dicos nunca buscan.
Los profesionales de la medicina funcional o los m�dicos observan los niveles de amino�cidos, niveles de minerales e incluso los niveles de toxinas de los metales pesados ??como mercurio, plomo, ars�nico y muchos m�s. Tambi�n analizamos sus niveles de antioxidantes, niveles de vitamina A y vitamina E, as� como su antioxidante CoQ10 y el estado de betacaroteno. Podemos determinar si una persona come verduras o no si, a modo de ejemplo, tienen niveles bajos de betacaroteno. Tambi�n analizamos los niveles de vitamina D, �cidos grasos esenciales, incluidas las grasas omega-3 y las grasas omega-6. Podemos determinar si una persona come comida chatarra. Podemos determinar si una persona est� comiendo pescado. Y podemos determinar si una persona est� comiendo demasiado aceite de oliva o grasas saturadas. Todo est� demostrado en estas medidas y pruebas.
Una prueba OAT, o prueba de �cidos org�nicos, tambi�n analiza lo que se conoce como �cidos org�nicos. Esta prueba demuestra una amplia gama de par�metros asociados con sus mitocondrias, que analizaremos en el siguiente art�culo, sus vitaminas B, sus neurotransmisores, su flora intestinal y su desintoxicaci�n. En �ltima instancia, es un examen completo que me muestra si un paciente est� bien o enfermo. Me muestra d�nde est�n los desequilibrios y d�nde debo recomendar modificaciones en su estilo de vida. Tambi�n ayuda a proporcionar pistas sobre otros problemas de salud.
A modo de ejemplo, si sus mitocondrias no funcionan correctamente porque tiene niveles reducidos de amino�cidos esenciales o si tiene un mayor estr�s oxidativo o si simplemente tiene niveles bajos de selenio y zinc, existe la posibilidad de que tenga alg�n tipo de sobrecarga t�xica por metales pesados. Y eso es precisamente lo que ir�a buscando. Los signos como estos proporcionan mucha informaci�n sobre lo que podemos hacer para tratar a un paciente. Y un m�dico o practicante de medicina funcional con experiencia puede determinar qu� est� pasando realmente con un paciente o puede ayudar a los pacientes a descubrir c�mo optimizar su salud y bienestar en general.
La nutrici�n es el estudio de los nutrientes en los alimentos y c�mo el cuerpo humano utiliza los nutrientes, as� como la relaci�n entre la dieta, las enfermedades, la salud y el bienestar en general. Los nutrientes son una fuente de nutrici�n, incluidos los carbohidratos, prote�nas, grasas, vitaminas, minerales, fibra y agua. La medicina funcional se enfoca en el uso de alimentos como una forma de medicina. Una nutrici�n equilibrada puede ayudar a prevenir y tratar una variedad de problemas de salud. De manera similar, la nutrici�n en la medicina funcional implica c�mo ciertas enfermedades y condiciones pueden estar asociadas con factores diet�ticos, como una mala alimentaci�n o malnutrici�n, alergias a los alimentos e intolerancias a los alimentos. Dr. Alex Jimenez D.C, C.C.S.T.
Entendiendo su Nutrici�n
Como buenos doctores de medicina funcional, a menudo nos preguntamos, �por qu� es que tantas personas en los Estados Unidos est�n sobrealimentadas pero desnutridas? O, �por qu� es que los estadounidenses comen demasiadas calor�as y muy pocos nutrientes? Las principales causas de las deficiencias nutricionales generalizadas son las siguientes: Primero, los humanos evolucionaron de comer alimentos silvestres que conten�an niveles tremendamente m�s altos de nutrientes. Segundo, el suelo que utilizamos actualmente para cultivar nuestros cultivos se ha agotado en gran medida de nutrientes. Las t�cnicas de hibridaci�n de la agricultura industrial est�n produciendo animales y vegetales para tener niveles reducidos de nutrientes. Tercero, los alimentos procesados ??no tienen absolutamente ning�n nutriente, por lo que con frecuencia tienen que ser fortificados. Y por �ltimo, pero no menos importante, la exposici�n a las toxinas ambientales, la falta de luz solar, el estr�s cr�nico y la mala alimentaci�n, incluido el aumento del consumo de alcohol, cafe�na y az�car, pueden aumentar nuestras necesidades nutricionales, muchas de las cuales ya no estamos obteniendo lo suficiente de nuestra dieta.
Bueno, es posible que no necesite ninguna vitamina, sin embargo, si puede cumplir con ciertas condiciones. Tal vez si solo cazara y recolectara alimentos silvestres y no estuviera expuesto a toxinas ambientales. O tal vez si se acostaba con el sol y se despertara con el sol, durmiendo nueve horas por noche. Y si no experimenta absolutamente ninguna cantidad de estr�s cr�nico. En �ltima instancia, si solo bebe agua pura y limpia y respira aire puro y limpio. Entonces, probablemente no necesitar�a vitaminas. Pero el resto de nosotros que no seguimos estas condiciones, si las necesitamos.
Y con ese pensamiento, terminamos este art�culo. En el pr�ximo art�culo, hablaremos de hormonas. Las hormonas pueden afectar casi todos los aspectos de nuestro bienestar, y muchos profesionales de la salud no entienden cu�les deben ser nuestros niveles hormonales �ptimos o incluso cu�ndo probarlos y qu� hacer al respecto una vez que lo hacen. La medida y prueba de los niveles hormonales debe ser una pr�ctica est�ndar, y muchos pacientes nunca han tenido un panel de sangre para observar sus hormonas. Es fundamental saber y comprender lo que sucede dentro de su propio cuerpo. Y es por eso es que este pr�ximo art�culo es tan importante. No querr� perderse de nuestra pr�xima actualizaci�n. Los veo pronto. El alcance de nuestra informaci�n se limita a problemas quiropr�cticos y de salud de la columna, as� como a temas y discusiones de medicina funcional. Para seguir discutiendo el tema, no dude en preguntarle al Dr. Alex Jimenez o comun�quese con nosotros al�915-850-0900�.
Curado por el Dr. Alex Jim�nez
Discusi�n del Tema Adicional: Dolor de Espalda Agudo
El dolor de espalda es una de las causas m�s frecuentes de discapacidad y d�as perdidos en el trabajo en todo el mundo. El dolor de espalda se atribuye a la segunda raz�n m�s com�n para las visitas al consultorio del m�dico, superada �nicamente por infecciones respiratorias superiores. Aproximadamente el 80 por ciento de la poblaci�n experimentar� dolor de espalda al menos una vez a lo largo de su vida. La columna vertebral es una estructura compleja formada por huesos, articulaciones, ligamentos y m�sculos, entre otros tejidos blandos. Las lesiones y / o afecciones agravadas, como las hernias de disco, pueden provocar s�ntomas de dolor de espalda. Las lesiones deportivas o las lesiones por accidentes automovil�sticos suelen ser la causa m�s frecuente de dolor de espalda; sin embargo, a veces los movimientos m�s simples pueden tener resultados dolorosos. Afortunadamente, las opciones de tratamiento alternativo, como la atenci�n quiropr�ctica, pueden ayudar a aliviar el dolor de espalda mediante el uso de ajustes de la columna vertebral y manipulaciones manuales, lo que finalmente mejora el alivio del dolor. �
De XYMOGEN�Las f�rmulas profesionales exclusivas est�n disponibles a trav�s de profesionales de atenci�n m�dica con licencia seleccionados. La venta por internet y el descuento de f�rmulas XYMOGEN est�n estrictamente prohibidos.
Con orgullo�El Dr. Alexander Jimenez�hace que las f�rmulas de XYMOGEN est�n disponibles solo para los pacientes bajo nuestro cuidado.
Llame a nuestro consultorio para que podamos asignar una consulta m�dica para acceso inmediato.
Si eres paciente de�Injury Medical & Chiropractic�Clinic, puedes preguntar por XYMOGEN llamando�915-850-0900.
Para su conveniencia y revisi�n de la�XYMOGEN�productos por favor revise el siguiente enlace. *XYMOGEN-Descargar-Catalogo
* Todas las pol�ticas de XYMOGEN anteriores se mantienen estrictamente.
IFM's Find A Practitioner tool is the largest referral network in Functional Medicine, created to help patients locate Functional Medicine practitioners anywhere in the world. IFM Certified Practitioners are listed first in the search results, given their extensive education in Functional Medicine