Calcium overview- Calcium is a mineral that is essential for life (2). Not only does the body require calcium to build strong bones, but it also aids the body in keeping the bones strong, muscle contractions, and helps to enable our blood to clot (2). Majority of the calcium in the body can be found in the bones and teeth. However, we lose calcium every day just through our skin, nails, hair, and sweat. Once calcium is dissolved in the stomach, it is then absorbed through the small intestine lining to enter the bloodstream. From here, the calcium can then build bone and regulate the contraction of the blood vessels as well as perform its other duties (2).� One of the reasons the amount of calcium consumed each day is so important is because, no matter what, the body will take what it needs. This being said, if you are not supplying your body with the correct amount of calcium, it will start to take the nutrients it needs from the bones (1). The more and more the body does this, the more fragile your bones become, leaving you more susceptible to diseases such as osteoporosis. However, there are different forms of calcium that provide and aid the body in different things.
Calcium D Glucerate
Calcium D Glucerate is made in small amounts by humans. This is the calcium salt of D Glucerate (1). With studies performed, results showed that when Calcium D Glucerate is taken orally, it inhibits beta-glucuronidase (1). When beta-glucuronidase is inhibited, it aids the body in preventing many hormone-dependent cancers. These cancers include but are not limited to; breast, prostate, and colon (1). When beta-glucuronidase is elevated in the body,� cell damage begins to occur. However, when Calcium D Glucerate is taken orally, it helps to inhibit (block) this enzyme that is produced by the liver.
Calcium Carbonate
Calcium Carbonate is calcium with a salt formula. This medicine can be used in multiple situations but is most commonly used for temporary relief of heartburn and indigestion (3). In addition to this, Calcium Carbonate can be used to help prevent osteoporosis (3).
The recommended daily dose for adults is 1,000mg a day of calcium. Make sure that you are eating foods containing calcium as well as taking this recommended dose in order to best protect your bones. Not only will this help your bones from becoming porous, but it will aid in overall body performance. If you are confused about which calcium supplement you should be taking, please consult a local doctor. – Kenna Vaughn, Senior Health Coach
Sources:
�Calcium-D-Glucarate.� Alternative Medicine Review: a Journal of Clinical Therapeutic, U.S. National Library of Medicine, Aug. 2002, www.ncbi.nlm.nih.gov/pubmed/12197785.
�Calcium/Vitamin D Requirements, Recommended Foods & Supplements.� National Osteoporosis Foundation, 26 Feb. 2018, www.nof.org/patients/treatment/calciumvitamin-d/.
National Center for Biotechnology Information. PubChem Database. Calcium carbonate, CID=10112, https://pubchem.ncbi.nlm.nih.gov/compound/Calcium-carbonate (accessed on Aug. 11, 2010
Supplemental Facts title indicates that the product is marketed for sale in the USA and is FDA standard.
Serving size, and the number of servings per container will be included to help you compare between products.
Make sure the serving sizes match when comparing supplements to get an accurate comparison.
Vitaminsand minerals always show the dose in weight and % percentage daily value to help you understand your dietary requirements.
Supplements often will have doses that exceed the recommended daily value.
Dietary supplement ingredients that are not vitamins or minerals will not have the % percentage daily value because they are not essential ingredients in the diet.
Dietary supplements are regulated by the FDA, and all labels must follow a consistent format to make it easier for consumers to understand. Understanding dietary supplement labels well require some attention to the following points when you�re evaluating supplements.
Herbs sometimes have additional information listed in the supplement panel. You might see ratio numbers (example 4:1) that show how much raw material of the herb (fresh or dried herb) is in the supplement version.
Herbs might have a standardization amount that shows how much of an active ingredient is present. The dose of the active ingredient is often listed, but sometimes it is not.
Proprietary blends are common. Only the total amount of the blend in a serving needs to be listed, which means that you don�t get all of the information about every ingredient.
Proprietary blend ingredients are listed in order from most to least. Similar to how food ingredients are listed on prepared foods nutrition facts panels.�
The daily value percent is established against a 2000 calorie diet. This is the standard calorie amount, however, it�s always important to determine your calorie requirements that you need for your health goals.
How To Identify Organic Foods
In the United States, a food or product that is labeled as organic is required to be certified by the�U.S. Department of Agriculture (USDA). The USDA has a certification program for natural growers and it has a set of very stringent standards that the product or food must meet.
There are some exemptions. For instance, a producer who does not sell more than $5,000 annually just in organic foods is not required to get the certification although they do have to adhere to the USDA�s stringent requirements for organic foods.
When a food carries the USDA Organic label, it means that it meets the requirements. While natural producers are not required to put the label on their products, many do.
The labeling varies, depending on the type of food. Single-ingredient foods like eggs, vegetables, and fruits�are considered to be 100 percent natural and are allowed to carry the USDA seal.
Foods that contain two or more ingredients, like breakfast cereal, are still allowed to use the USDA seal, but also must include the following information:
Organic � The product must be 95 percent organic or greater in order to be able to use this term
100 percent organic � The product must be completely organic or all of its ingredients must be natural
Made with organic ingredients � The product contains no less than 70 percent natural ingredients
If the product has less than 70 percent natural ingredients, they are not allowed to use the word �organic� anywhere on their product labels.
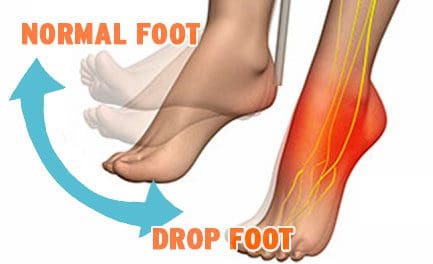
Drop Foot
Drop Foot is a symptom associated with an underlying neurological, muscular or anatomical problem that is often treated with a combination of non-invasive techniques and procedures.
Overview
Drop foot/foot drop is a general term that means lifting the forefoot (front part of the foot) is difficult to do.
Drop foot is not a diagnosis but a symptom associated with an underlying condition such as:
Neurological
Muscular
Anatomical problem
Nerve or muscle injury
Brain/spinal cord disorder, including
Herniated disc
Spinal Stenosis
Spondylolisthesis
Vertebral Fractures
Walking can be difficult, so the foot tends to drag on the floor.
Those suffering from drop foot sometimes raise the thigh up in an exaggerated fashion, such as when climbing stairs.
Stepping high is the most common symptom.
Symptoms of Drop Foot
High steppage gait
Foot drop may be experienced in one or both feet.
It is typically experienced in one foot if the drop foot is caused by:
Braces can provide additional support, stability, and shock absorption
Specific exercises to help the affected muscles
Physical therapy
Prevention
Prevention includes keeping your bones and tissues strong and healthy through diet and�exercise.
Avoid tobacco and excessive alcohol use creates weak bones and problems with the blood supply
Reduce the risk of injury by following safety measures on the job and beyond (i.e. wear a seatbelt).
Custom orthotics can provide additional support, stability, and shock absorption.
Kneeling for extended periods, such as on the job (certain construction functions laying tile, yard work).
Recovery
Some cases of drop foot are temporary; however, others can become permanent.
The recovery time and process depend on the cause.
Reduce *PLANTAR FASCIITIS PAIN* with Custom Foot Orthotics | El Paso, TX (2019)
Foot problems can affect the overall well-being of the human body. Many health issues which affect the foot can result in poor posture, low back pain, and sciatica. These imbalances can ultimately result in a variety of other health issues. Custom-made foot orthotics can help promote and support overall well-being by relieving foot problems.
Dr. Alex Jimenez is the non-surgical choice for foot problems and other health issues. Dr. Alex Jimenez can help promote and support overall well-being with the use of custom-made foot orthotics and other treatments approaches.
What’s Afoot
Foot Dysfunction can very easily cause a domino effect that extends all the way to the back. The feet are the foundation of the body and when there is a problem with the way they function it can cause the entire body to shift out of alignment. Overpronation and oversupination, for example, can cause a variety of injuries and conditions that affect not only the feet and ankles, but also the knees, hips, and back as well.
NCBI Resources
If you have further questions or concerns about your particular�diet, please ask us! Our Doctor of Chiropractic can help guide you toward a more healthy life, including the foods you consume. It comes down to two major areas: safety and�nutrition. That is what consumers need to understand when they are trying to make a decision on whether to purchase foods that have been conventionally farmed or foods that are natural.
Supplements can become overwhelming with everything that’s out there. This will touch on key categories of supplements and the best way on how to use them.
Supplements are divided into categories:
Nutrients
(Nutraceuticals or Orthomolecular Medicine)
Botanicals
(Herbal Medicine or Phytotherapy)
Hormones
Hormone Replacement Therapy or Restorative Medicine
Each category is meant to support underlying excesses or deficiencies, with the ultimate goal of bringing the body back into balance.
Where it becomes complicated is understanding the different uses and dosing of specific:
Extracts
Chelates (a chemical compound that dissolves more easily and is easily absorbed compared to other types of molecules and compounds)
Prepared Compounds
Hormones
Integrative medicine and hormone therapy usually involve synthetic versions of plant extracts, that mimick the chemical structure of endogenous hormones.
What are Hormones?
Hormones are chemical messengers that are produced by the endocrine glands.
These messengers have an effect on most tissues and major bodily functions.
Establishing and resolving the causes of hormonal imbalance is one of the core therapeutic systems not only in integrative medicine but in conventional medicine.
How do Hormones Work?
Hormones flow through the body�s circulation in abundance. In order for hormones to perform a function, the cells being used require specific receptors.
When hormones bind to these receptors, it causes a biological response inside the cell, which is why a single hormone can have such an effect.
The entire endocrine system is based on positive and negative feedback loops. This informs the endocrine glands to produce more or fewer hormones.
Balance of endogenous hormone production is maintained through diet and lifestyle. However,�when this unique balance is disrupted is when disease can occur.
Despite their broad and potent effect on the body’s systems, hormones therapy must be done with caution and skill.
There can be long-term negative side effects with hormone therapy and should be limited to highly-trained practitioners.
Examples of Hormone Ingredients
Exogenous hormones are available in different forms.
Examples include:
Pregnenolone – this is the mother of steroidogenic hormones
Dehydroepiandrosterone (DHEA) – elevates estrogen and testosterone
Vitamin D – regulates bone density, and balances parathyroid hormone secretion
There is evidence that shows beneficial therapeutic uses for exogenous hormones.
Similar to botanical medicine, hormone therapy should be used exclusively and with caution by certified clinicians.
Fasting Mimicking Diet Basics
Some people today might eat a slightly higher amount of calories the first day as they ease into the fasting-mimicking diet. They might then decrease their total caloric intake. You also want to make sure you eat smaller amounts of foods which are easy to digest.
ProLon��offers a pre-packaged box which contains all five days� worth of meals for you to do the diet. The meals are all plant-based. One day, by way of instance, offers tea and a nut bar for breakfast, a small portion of vegetable soup and a few kale crackers for lunch, several olives in the afternoon, and finally another small portion of vegetable soup for dinner.
You can also do the fasting-mimicking diet without the need for a pre-packaged box-like ProLon�. Simply follow the right proportions and plan out how you will space them out every day. Macros for the fast mimicking diet are 34 percent carbohydrates, 10 percent protein, and 56 percent fat for the very first day and 47 percent carbohydrates, 9 percent protein, and 44 percent fat to the rest days.
A cup of black tea and coffee every day are generally allowed. Just make sure they don�t contain any added sugars or oils. Remember that people with health issues should consult a healthcare professional prior to doing the fast mimicking diet in your own home.
Foods
Dr. Anthony Gusting followed a four-day ketogenic fasting-mimicking diet. Every day, he consumed different amounts of bone broth, coconut milk, coconut oil, BCAAs, and exogenous ketones. Avocados and grass-fed butter can also be included in the fast mimicking diet. This is a great way to combine the ketogenic diet with the fasting-mimicking diet to benefit from the two regimens.
Supplements
Taking nutritional supplements can also make the fasting-mimicking diet easier by providing enough nutrition. These may include:
Electrolytes like magnesium and salt to replenish any lost during water loss
Grass-fed liver tablets to provide micronutrient support
Branch chain amino acids, or BCAAs, to help prevent loss of lean tissue
Greens powder to provide micronutrients
Algal oil or cod liver oil for omega-3s
You may also take exogenous ketones to achieve ketosis through the keto diet. The fast mimicking diet can also help you achieve ketosis before following a ketogenic diet. Below, we will discuss how the fast mimicking diet promotes ketosis.
Ketosis And The Fast Mimicking Diet
The fast mimicking diet is an excellent way to prepare you for the ketogenic diet. This is because it allows you to get into ketosis. Additionally, eating keto foods makes it possible to remain in ketosis throughout the regimen. To follow a ketogenic fasting-mimicking diet you must maintain your macros over the suitable range of 5 to 10 percent of carbohydrates, 20 to 25percent of proteins, and 70 to 80 percent of fats. If you�re unsure about whether you�re properly maintaining your macros, always choose something with more fat.
If you or a loved one need additional dietary guidance, give us a call. Our Doctor of Chiropractic is here to help!
*CORRECT BAD POSTURE* with Custom Foot Orthotics | El Paso, TX (2019)
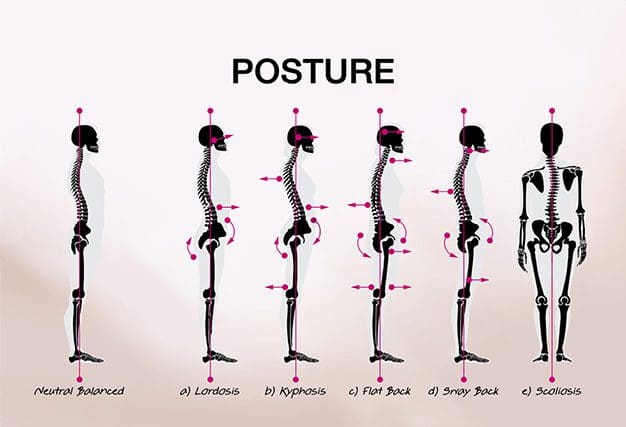
Posture is the way in which we hold our bodies while standing, sitting, or performing tasks such as lifting, bending, pulling, or reaching. However, when a person practices bad posture, it can cause a variety of health issues. According to research studies, signs and symptoms associated with a bad posture are the 3rd most common reason for doctor office visits in the United States.
The spine helps support the weight of the head and stabilizes the human body when standing upright. However, bad posture can strain the spine, causing a variety of health issues. Custom orthotics can help support good posture and promote pain relief. Dr. Alex Jimenez can help patients improve their overall health and wellness through custom orthotics. Patients recommend Dr. Alex Jimenez as the non-surgical choice for posture.
Posture
Poor posture takes a serious toll on health. Here are a few negative results:
Muscle soreness
Subluxations
Blood vessel constriction
Nerve constriction
As time passes, body/health problems also arise, which result in pain, restricted motion, and make the body more susceptible to injury and disease. Posture can even affect your emotional well-being, including self-confidence!
Treatment
Chiropractic can help you maintain proper posture with alignment adjustments, and custom orthotics can benefit the treatment making it last longer.
The American Chiropractic Association maintains proper posture involves sitting, standing, and lying down properly. Here’s an excerpt:
Sitting properly
Keep your feet on the floor or footrest, for feet to be stabilized.
Ankles should be in front of your knees.
Keep a small space between the back of the knees and the front of the seat.
Your knees should be at or below the level of your hips.
Standing properly
Keep your weight on the balls of your feet.
Keep your knees slightly bent.
Keep your feet in line with your shoulders.
Arms hang naturally down the sides of the body.
Sleeping properly
Find the mattress that is right for you.
A firm mattress is generally recommended but sometimes people do better with a
Softer mattresses to reduce back pain.
Remember comfort is important.
Sleep with a pillow.
Special pillows are available to help with posture problems that come from a poor sleeping position.
The feet are complex structures which function to provide protection and support to the body in relation to the spine and head posture. The feet are essential towards balancing the body�s entire weight, allowing it to walk, run, stand, and, jump. Foot complications may cause severe issues throughout the spine, hips, knees, and ankles if left untreated. Chiropractic care can be effective for treating injuries or conditions such as plantar fasciitis, Achilles tendonitis, and ankle sprains. Foot health is essential towards establishing the body�s proper support and balance, together with a proper spinal posture. In the presence of foot complications, chiropractic treatment can be used to achieve long-lasting health.
NCBI Resources
It seems our moms were right; we are what we eat. Unfortunately, in today�s marketplace, there are many valid worries about the foods we consume. Antibiotics, pesticides, and foods tainted with dangerous diseases are a top concern for many Americans. In addition, processed and genetically modified foods�and artificial ingredients contribute to a number of health issues from�obesity�to�cancer.
Supplements can become overwhelming with everything that’s out there. This will touch on key categories of supplements and the best way on how to use them.
Supplements are divided into categories:
Nutrients
(Nutraceuticals or Orthomolecular Medicine)
Botanicals
(Herbal Medicine or Phytotherapy)
Hormones
Hormone Replacement Therapy or Restorative Medicine
Each category is meant to support underlying excesses or deficiencies, with the ultimate goal of bringing the body back into balance.
Where it becomes complicated is understanding the different uses and dosing of specific:
Extracts
Chelates (a chemical compound that dissolves more easily and is easily absorbed compared to other types of molecules and compounds)
Prepared Compounds
Botanicals
(Herbal Medicine or Phytotherapy)
Botanicals?
Records of humanity using botanical extracts therapeutically date back to the beginning of first recorded history.
Today, botanical medicine serves as an inspiration for pharmaceutical inventions and continues�via modern research.
Botanical medicine is essential in integrative medicine and normally elicits fewer and less significant adverse side effects than pharmaceutical interventions.
Growing research shows that when used correctly, botanical extracts are more effective than pharmaceutical equivalents, and more cost-effective as well.
Botanical Work?
Given their capacity to interact with virtually every system and tissue in the body, there are thousands of therapeutic uses.
There is a great deal of literature on traditional botanical medicine, however, there is a very little amount of modern research relative to modern medicine.
As a result, it becomes somewhat challenging to compare botanicals with pharmaceuticals regarding
clinical outcomes.
Botanical Ingredients
Botanical medicine provides very helpful effective therapeutics and allows everyone, from professional practitioners to common laypeople apply these treatments.
Common examples:
Garlic (Allium sativum) – immune function, cardiovascular function
Bottom line, diets are temporary. There are some serious consequences that can come from dieting, especially fad or crash diets. The effects of these types of diets can also seriously impact your chiropractic care, hindering your progress.
Also, because diets are temporary, once you return to your regular eating habits the weight usually comes back.
A lifestyle change is a far better choice. It involves making smart, healthy eating choices � choices that you maintain for the rest of your life. This also impacts your chiropractic care by strengthening your body and keeping it healthy so that it is in an optimal state for healing and responds well to treatment.
Diet Type
There are all sorts of diets out there. Some are blatantly unhealthy but others are sneaky. They come with claims of being healthy, of being created or endorsed by doctors, or include tons of vitamin supplements but very limited food intake. It is important to be able to spot these destructive fad diets so you don�t get suckered into their hype.
Some of the most common�types of fad diets include high protein, low or no-carb, liquid, cabbage, grapefruit, broth or juice, and food combining. Some of these can cause serious health problems including vital organ damage. Others can cause vitamin deficiencies and dehydration. None of them can (or should) be maintained over a long period of time, much less the rest of your life.
Healthy Eating/Lifestyle
When you make the life-changing commitment to adopt a healthy eating lifestyle you open yourself up to a world of better health, more energy, and better focus. Your body will heal faster and you will feel better.
A diet of fresh�fruits and vegetables, lean meats, fresh fish, and whole grains, along with lots of water should become a way of life. It is far healthier than the temporary diets that are out there and more effective too.
If you or a loved one need additional dietary guidance, give us a call. Our Doctor of Chiropractic is here to help!
Get Rid of *FOOT PRONATION PAIN* with Custom Foot Orthotics | El Paso, TX (2019)
Foot pronation is the natural movement which occurs during foot landing while walking or running. Foot pronation also occurs while standing, and in this instance, it is the amount in which the foot rolls inward toward the arch. Foot pronation is normal, however, excessive foot pronation can cause a variety of health issues, including bad posture. The following video describes the 5 red flags of excessive foot pronation, which can ultimately affect a person’s overall health and wellness. Dr. Alex Jimenez can help diagnose and treat excessive foot pronation. Patients recommend Dr. Alex Jimenez and his staff as the non-surgical choice for excessive foot pronation health issues.
What’s Afoot
The feet are complex structures which function to provide protection and support to the body in relation to the spine and head posture. The feet are essential towards balancing the body�s entire weight, allowing it to walk, run, stand, and, jump. Consequently, foot complications may cause severe issues throughout the spine, hips, knees, and ankles if left untreated. Chiropractic care can be effective for treating injuries or conditions such as plantar fasciitis, Achilles tendonitis, and ankle sprains, among others. Foot health is essential towards establishing the body�s proper support and balance, together with a proper spinal posture. In the presence of foot complications, chiropractic treatment can be used to achieve long-lasting health.
NCBI Resources
While most of us know that proper nutrition is vital in assisting us to reach our optimum health and feel our best; finding time to eat a balanced diet on a daily basis seems a formidable job in this fast-paced, affluent society. Though your life may be frantic, there are still many good tasting, healthful alternatives which can assist you to lose weight and enhance your health.
Supplements can become overwhelming with everything that’s out there. This will touch on key categories of supplements and the best way on how to use them.
Supplements are divided into categories:
Nutrients
(Nutraceuticals or Orthomolecular Medicine)
Botanicals
(Herbal Medicine or Phytotherapy)
Hormones
Hormone Replacement Therapy or Restorative Medicine
Each category is meant to support underlying excesses or deficiencies, with the ultimate goal of bringing the body back into balance.
Where it becomes complicated is understanding the different uses and dosing of specific:
Extracts
Chelates (a chemical compound that dissolves more easily and is easily absorbed compared to other types of molecules and compounds)
Prepared Compounds
Nutrients
Nutrients are defined as ingredients that would otherwise be found in food, which include:
Vitamins
Minerals
Phytochemicals
Antioxidants
What are Nutrients?
For sustaining life and staying healthy, the human body requires a balance of nutrients, which can be categorized as macronutrients and micronutrients.
Macronutrients constitute the major food groups of
Carbohydrates
Fats
Proteins
These are required in large amounts and are used for energy production and tissue growth.
Micronutrients are required in small amounts and include vitamins and minerals.
Phytonutrients, such as anthocyanin found in blueberries or resveratrol found in grapes, are also often classified as micronutrients.
How do Nutrients Work?
Macronutrients are the caloric basis for our
Daily energy needs
Provide the building blocks of tissue development
Fiber for our microbiota
Cholesterol for hormone production
Micronutrients are not relevant for energy and contribute to:
Tissue development
Hormone and Neurotransmitter Production
Brain function
Immune function
Common Nutritional Supplements
Dietary supplements can provide nutrients in a condensed form, which allow for targeting specific conditions or in order to upregulate certain pathways that require those nutrients.
Examples include:
Whey Protein Isolate
Omega 3 Fatty Acids
Calcium
B vitamins
Eating Properly/Healthy & Staying Hydrated
Making Protein A Priority
Muscle injuries are one of the most popular reasons for chiropractic visits. Protein helps build and heal muscle tissue, helping it to renew itself back to a pre-injury state.
If your daily intake of protein is low, it can hinder the healing process and stand in the way of chiropractic visits giving you the maximum results. Fish and lean meats are the foods that offer the highest protein.
Other vegetarian options are tofu, soybeans, eggs, milk, and nuts. Help your muscles rebuild and heal by adding high protein foods into every meal menu.
Staying H2o Hydrated
Why do most of us view water drinking with dread? If you wish to promote healing, it�s imperative to drink enough water every day.
Water provides an entire slew of benefits, from transporting those important nutrients to where they need to be, to flushing harmful toxins out of the body. Aim for eight glasses of water a day, and try to drink it throughout the day.
Getting into this habit will boost overall health, and promote healing of injuries and other medical conditions.
While it�s a smart goal to try to ingest all the nutrients you need through food and drink, supplements are available to bridge the gap. Before beginning a supplement regimen, talk to you chiropractor for recommendations on the type and dosage that will aid and improve your unique situation.
You may also want to download a smartphone app to track your daily intake of calories, calcium, protein, and liquid. There are many available, one being My Fitness Pal.
Chiropractic treatment is a big step in the right direction for overall health/well-being and incorporating these nutritional tips into your daily routine will help you get the most out of your chiropractic care.
Correct your Bad Posture with *FOOT ORTHOTICS* | El Paso, Tx
Posture is the position in which we hold our bodies while standing, sitting, or lying down. Good posture allows the body parts to be properly supported through correct alignment to promote the right amount of tension against gravity. Bad posture can cause a variety of health issues, including neck pain and back pain. Understanding how bad posture can affect your health is essential for overall wellness. Custom foot orthotics can also help improve posture. Dr. Jimenez is the non-surgical choice for posture health issues.
What’s Afoot
Treatment for foot pain varies depending on the condition/injury. Treatment can go from rest and ice to physical therapy, chiropractic and in severe cases surgery. Reflexology can provide relief, as well as, stretching exercises. Over the counter pain medication is often used. If the pain is too intense that it prevents sleep, a physician may prescribe non-addictive pain medication. Wear shoes with good arch supports, and if pain persists, see a podiatrist for special orthotic shoe inserts. Insurance often covers orthotics.
NCBI Resources
Don�t forget that most pain in the body is caused by inflammation and can be helped with anti-inflammatory diet and lifestyle stressors. Concentrate on eating whole, unprocessed foods. Stay away from sugar, alcohol, artificial sweeteners, and white flour. Make sure to drink enough water every day, and get eight hours of sleep. This is one of the most effective ways to address inflammation. Bring the body back into balance.
Magnesium is an essential mineral, yet it tends to be overlooked as a health concern. To illustrate, a study shows that up to 75% of individuals are not intaking the recommended daily amount (2). This stems from the fact that most foods have steadily decreased the amount of magnesium they provide. Thus leaving many consumers with the need to supplement. Different magnesium supplements have been shown to facilitate the body in different ways.� Magnesium citrate can help to lower blood pressure whereas Magnesium L-Threonate can help with memory loss. So, which magnesium supplement should you be taking?��� �
Magnesium Citrate Overview
Magnesium Citrate is one of the most bioavailable forms of magnesium out there, meaning it’s ready and easily absorbed by the body’s digestive tract (5). In addition to this, Magnesium Citrate has been shown to improve arterial stiffness, lower blood pressure, and in turn, help to prevent cardiovascular disease in patients who are slightly obese (3). However, be aware that magnesium citrate is also used to treat constipation, therefore it can act as a laxative by helping to absorb water in the intestines (1). �
Magnesium L-Threonate Overview
Magnesium L-Threonate is a newer form of magnesium, therefore there are fewer studies showing its full capabilities. However, Magnesium L-Threonate is the only form of magnesium to cross the blood-brain barrier more readily. The blood-brain barrier is extremely selective in order to reduce the amount of toxins entered into the CNS (central nervous system). Due to this, it has been proven to increase synapse density and aid in memory loss, cognitive decline, as well as help to improve short term memory (4). In a long term study of 17 years performed with Magnesium L-Threonate, the results manifested that subjects were 37% less likely to get dementia (4). �
If you are looking for something to assist in relaxation, sleep, and memory; Magnesium L-Threonate is the supplement for you. Although, if you are looking for something to relieve occasional constipation and something to aid in blood pressure that mixes well with water and has little to no taste, magnesium citrate is the route you should consider. Getting started on the right magnesium supplement today could set you up with a better tomorrow. – Kenna Vaughn, Health Coach Insight
The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues as well as functional medicine articles, topics, and discussions. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 . �
References
(1)Cisar�, Fabio, et al. �Bowel Preparation for Gastrointestinal Endoscopic Procedures With Sodium Picosulphate-Magnesium Citrate Is an Effective, Safe, and Well-Tolerated Option in Pediatric Patients: A Single-Center Experience.� Gastroenterology Nursing : the Official Journal of the Society of Gastroenterology Nurses and Associates, U.S. National Library of Medicine, 2018, www.ncbi.nlm.nih.gov/pubmed/30063687.
(2)Guerrera, Mary P, et al. �Therapeutic Uses of Magnesium.� American Family Physician, U.S. National Library of Medicine, 15 July 2009, www.ncbi.nlm.nih.gov/pubmed/19621856.
(3)Schutten, Jo�lle C, et al. �Effects of Magnesium Citrate, Magnesium Oxide and Magnesium Sulfate Supplementation on Arterial Stiffness in Healthy Overweight Individuals: a Study Protocol for a Randomized Controlled Trial.� Trials, BioMed Central, 28 May 2019, www.ncbi.nlm.nih.gov/pubmed/31138315.
(4)”Science Review: Magnesium L-Threonate.” Metagenics Institute. 01 Aug. 2019 .
(5)Walker, Ann F, et al. �Mg Citrate Found More Bioavailable than Other Mg Preparations in a Randomised, Double-Blind Study.� Magnesium Research, U.S. National Library of Medicine, Sept. 2003, www.ncbi.nlm.nih.gov/pubmed/14596323. �
Orthotics Treat Other Areas Than the Feet
�
�
Additional Topic Discussion: Xymogen� Supplements
The average adult in the United States takes one or more dietary supplements on a regular basis. Dietary supplements include vitamins, minerals, herbals and botanicals, amino acids, enzymes, and a variety of other products. Several of the most common dietary supplements include vitamin D and vitamin E, minerals such as calcium and iron, herbs such as echinacea and garlic, and products such as glucosamine, probiotics, and fish oils. For people who don’t have balanced nutrition, taking dietary supplements can help them get necessary amounts of essential nutrients and can help improve overall well-being.
Formulas for Methylation Support
�
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
� Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
� Please call our office in order for us to assign a doctor consultation for immediate access.
� If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download � * All of the above XYMOGEN policies remain strictly in force. �
Can methylation donors help promote a balanced methylation support?
Many doctors and functional medicine practitioners generally recommend higher doses of methyl donors, such as 5-methyltetrahydrofolate, or 5-MTHF, and methylcobalamin, in several patients. By way of instance, people with genetic polymorphisms and people with out-of-range methylation-related biomarkers, such as in hyperhomocysteinemia, may often develop health issues which may affect the function of specific enzymes, such as methylenetetrahydrofolate reductase, or MTHFR, among others.
Nutrient deficiencies associated with methyl donors are a prevalent finding in laboratory evaluations, and depending on your population, these may be closely associated with vitamin B12 deficiency-related neuropathy, which is relatively common. Many doctors and functional medicine practitioners also understand the importance of methylation support to help improve hereditary or environmental epigenetic health issues. Improving methylation status and avoiding the pathways of disease and dysfunction associated with potential nutritional deficits in methylation activity is the goal of many healthcare professionals.
However, as with any other biochemical process, methylation activity occurs through the balance of homeodynamics, or the dynamic form of homeostasis. An imbalance in these biochemical processes can ultimately lead to dysfunction and disease. Therefore, although we can be confident that making sure we have enough methyl donors available for use in the human body is essential, we have to question whether �pushing� reaction rates utilizing supraphysiological doses are safe and effective. Instead of forcing reaction rates, perhaps the goal of healthcare professionals should ultimately be to allow the human body to function correctly.
Methylation Supplementation
Numerous health issues associated with long-term, high-dose supplementation for methylation donors include:
According to research studies, the effects of these genetic alterations remains unclear. In comparison to the altered function of MTHFR C677T and A1298C single nucleotide polymorphisms, or SNPs, which have been moderately evaluated, the diagnosis of other SNPs can demonstrate the overall function of these specific enzymes. The effect of these SNPs on methylation activity depends on enzymes working together in the circumstance of a person’s internal and external environment. These outcome measures have been demonstrated in a variety of genome research studies. As a result, researchers are unable to determine the effects which these alterations, including that of MTHFR C677T, can develop on a patient’s overall methylation status.
The correct supplementation dose for methylation donors remains mostly unknown, as it may vary tremendously between patients. No research studies have currently demonstrated what the correct dosage or duration of methyl-donor supplementation is required to balance biochemical and epigenetic methylation status. Various side effects of high-dose 5-methyltetrahydrofolate, or 5-MTHF, supplementation have been demonstrated in clinical practice, including anxiety and worsening of symptoms.
Hypermethylation may be detrimental to an individual’s overall health and wellness. The following series of articles contain many examples of region-specific DNA hypermethylation health issues, including immune dysfunction, Downs Syndrome, and cancer. Both DNA hyper- and hypomethylation can develop due to deficiencies in methylation donors. Folic acid has been associated with increased immune hypersensitivity and cancer. The bottom line is that we don�t understand what effect long-term, high-dose methylation donors and supplementation can have on DNA methylation support. Further research studies are still required to show this effect.
Methylation is a biochemical process which involves the conversion of methylation activity for the function of a variety of processes in the human body. Methylation imbalances, however, can cause a variety of health issues. Methylation donors are a form of supplementation for methylation support. But, research studies have demonstrated that supplementation for methylation support may not be right for everyone. The best way to improve methylation is through proper nutrition, physical activity, and exercise, among other dietary and lifestyle modifications. The nutritional guidelines below can help safely and effectively improve methylation support, promoting overall health and wellness.
Dr. Alex Jimenez D.C., C.C.S.T. Insight
Nutrition for Methylation Support
Methylation status can often depend on a patient’s dietary and lifestyle factors. Dietary and lifestyle factors, including physical activity and exercise, stress, sleep, medicines, and toxin exposure, plays a role in methylation. Single interventions with high-dose nutrient supplementation may lack long-term effectiveness or these may not achieve the desired methylation support.
Dietary and lifestyle modifications are safe and effective, alternative treatment options for most individuals with methylation imbalances. This may be fundamental for certain vulnerable individuals, such as patients with active cancers. Aging is also known to be associated with decreased methylation activity, therefore, the Methylation Diet and Lifestyle, or MDL, program can be utilized as an anti-aging tool. Methylation status diagnosis and treatment is essential during preconception, pregnancy and the postnatal period.
The MDL program and supplementation can help promote overall health and wellness. A dietary and lifestyle treatment approach can also be utilized as a follow-up plan for people who need high-dose nutraceutical support. A variety of foods can promote methylation support. Dietary and lifestyle modifications have also been demonstrated to considerably improve methylation activity.
In the following articles, we will discuss dietary and lifestyle factors which can help support methylation. We will also discuss the basic biochemistry of methylation, the roles of methylation in the human body, how to evaluate methylation status, and the risks and benefits of methyl donor supplementation as well as the health issues associated with too little or too much methylation activity.
Smoothies and Juices for Methylation Support
While many healthcare professionals can recommend nutritional guidelines and lifestyle modifications, there are several alternative treatment options you can try for yourself at home. As described above, however, supplementation for methylation support should be correctly determined by a healthcare professional. Smoothies and juices are a fast and easy way to include all the necessary nutrients you need for methylation support in a single serving. The smoothies and juices below are part of the Methylation Diet Food Plan.
Sea Green Smoothie Servings: 1 Cook time: 5-10 minutes � 1/2 cup cantaloupe, cubed � 1/2 banana � 1 handful of kale or spinach � 1 handful of Swiss chard � 1/4 avocado � 2 teaspoons spirulina powder � 1 cup water � 3 or more ice cubes Blend all ingredients in a high-speed blender until completely smooth and enjoy!
Berry Bliss Smoothie Servings: 1 Cook time: 5-10 minutes � 1/2 cup blueberries (fresh or frozen, preferably wild) � 1 medium carrot, roughly chopped � 1 tablespoon ground flaxseed or chia seed � 1 tablespoons almonds � Water (to desired consistency) � Ice cubes (optional, may omit if using frozen blueberries) Blend all ingredients in a high-speed blender until smooth and creamy. Best served immediately!
Sweet and Spicy Juice Servings: 1 Cook time: 5-10 minutes � 1 cup honeydew melons � 3 cups spinach, rinsed � 3 cups Swiss chard, rinsed � 1 bunch cilantro (leaves and stems), rinsed � 1-inch knob of ginger, rinsed, peeled and chopped � 2-3 knobs whole turmeric root (optional), rinsed, peeled and chopped Juice all ingredients in a high-quality juicer. Best served immediately!
Ginger Greens Juice Servings: 1 Cook time: 5-10 minutes � 1 cup pineapple cubes � 1 apple, sliced � 1-inch knob of ginger, rinsed, peeled and chopped � 3 cups kale, rinsed and roughly chopped or ripped � 5 cups Swiss chard, rinsed and roughly chopped or ripped Juice all ingredients in a high-quality juicer. Best served immediately!
Zesty Beet Juice Servings: 1 Cook time: 5-10 minutes � 1 grapefruit, peeled and sliced � 1 apple, washed and sliced � 1 whole beet, and leaves if you have them, washed and sliced � 1-inch knob of ginger, rinsed, peeled and chopped Juice all ingredients in a high-quality juicer. Best served immediately!
Protein Power Smoothie Serving: 1 Cook time: 5 minutes � 1 scoop protein powder � 1 tablespoon ground flaxseed � 1/2 banana � 1 kiwi, peeled � 1/2 teaspoon cinnamon � Pinch of cardamom � Non-dairy milk or water, enough to achieve desired consistency Blend all ingredients in a high-powered blender until completely smooth. Best served immediately!
ProLon� Fasting Mimicking Diet
alanced methylation support can be achieved through proper nutrition. The ProLon� fasting mimicking diet offers a 5-day meal program which has been individually packed and labeled to serve the foods you need for the FMD in precise quantities and combinations. The meal program is made up of ready-to-eat or easy-to-prepare, plant-based foods, including bars, soups, snacks, supplements, a drink concentrate, and teas. The products are scientifically formulated and great tasting. Before starting the ProLon� fasting mimicking diet, 5-day meal program, please make sure to talk to a healthcare professional to find out if the FMD is right for you. The ProLon� fasting mimicking diet can help promote methylation support, among a variety of other healthy benefits.
Many doctors and functional medicine practitioners may recommend higher doses of methyl donors in several patients, however, further research studies are needed to determine the proper amount of methylation supplementation. The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues as well as functional medicine articles, topics, and discussions. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 .
Curated by Dr. Alex Jimenez
Additional Topic Discussion: Acute Back Pain
Back pain is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. Your spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief.
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly, Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download
* All the above XYMOGEN policies remain strictly in force.
IFM's Find A Practitioner tool is the largest referral network in Functional Medicine, created to help patients locate Functional Medicine practitioners anywhere in the world. IFM Certified Practitioners are listed first in the search results, given their extensive education in Functional Medicine




 Herbs sometimes have additional information listed in the supplement panel. You might see ratio numbers (example 4:1) that show how much raw material of the herb (fresh or dried herb) is in the supplement version.
Herbs sometimes have additional information listed in the supplement panel. You might see ratio numbers (example 4:1) that show how much raw material of the herb (fresh or dried herb) is in the supplement version.