Do you often feel low brain endurance for focus and concentration? Do you often crave sugar and sweets in the afternoon? Or do you feel energized after meals? Glucose, or sugar, is the main source of energy in the human body. And, because the human brain has so many nerve cells or neurons, it is one of the most energy-demanding organs, which utilizes about one-half of all the energy from glucose in the human body. Sugar is important but too much of it can also have its downsides. �
Brain functions, such as memory, thinking, and learning, are relatively associated with glucose levels and how efficiently the brain utilizes this essential energy fuel source. If there isn�t enough glucose, or sugar, in the brain, by way of instance, neurotransmitters, or the human brain�s chemical messengers, don’t develop properly and the communications between neurons can ultimately break down. Additionally, dysglycemia, a common health issue caused by abnormal blood glucose levels, can cause loss of energy for brain function and has also been associated with poor attention and cognitive function. �
�The human brain is dependent on sugar or glucose as its main energy fuel source,� stated Vera Novak, MD, Ph.D., an HMS associate professor of medicine at Beth Israel Deaconess Medical Center. �It just simply cannot be without it.� �
What is Dysglycemia?
As previously mentioned, brain structure and function, such as cognition, can be affected by dysglycemia, or blood glucose abnormalities, in older adults. Researchers conducted a cross-sectional and longitudinal cohort research study, analyzing the association of dysglycemia with brain health. The researchers found that dysglycemia is associated with an increased number of brain infarcts, white matter hyperintensities volume, and decreased total white matter, gray matter, and hippocampus volume cross-sectionally. According to the research study, there was also a decrease in gray matter volume longitudinally. Dysglycemia was ultimately associated with reduced language performance, speed, and visuospatial function. �
�Our results suggest that dysglycemia affects brain health in elderly survivors, evidenced by higher cerebrovascular disease, lower white, and gray matter volume as well as language, visuospatial function, and cognitive speed,� stated the authors. �
Dysglycemia can cause changes in blood glucose levels which may cause a variety of health issues. Dysglycemia is also not necessarily defined by specific blood sugar levels. Instead, having an abnormally low, high, or unstable blood glucose levels suggests an underlying health issue that requires further investigation. Moreover, while type 1 and type 2 diabetes are the most common causes of dysglycemia, other examples of blood sugar level abnormalities can include gestational diabetes and pre-diabetic conditions as well as drug-related and genetically related abnormalities of the blood sugar levels. �
Furthermore, dysglycemia can be a result of hereditary or environmental factors, or it can even be a combination of both. Genes can predispose a person to ultimately develop dysglycemia over time, just as much as several lifestyle habits can, too. A poor diet high in unhealthy fats, sugars, and processed foods can commonly cause a person to develop dysglycemia. Lacking certain vitamins and minerals that enhance the human body�s sensitivity to insulin can also cause dysglycemia. �
Dysglycemia and Brain Health
Although the brain needs glucose or sugar, too much of this energy fuel source can also have several side-effects. A 2012 research study on animals conducted by researchers at the University of California at Los Angeles demonstrated a positive relationship between the consumption of fructose, another form of sugar, and the aging of cells. A 2009 research study, also utilizing animal models and conducted by a team of scientists at the University of Montreal and Boston College, connected excess glucose consumption to memory and cognitive deficiencies. Further research studies are still required. �
The effects of glucose and other forms of sugar on the human brain may be the most profound in diabetes, a group of health issues in which high blood glucose levels persist over a prolonged period of time. Type 1 diabetes is a health issue in which the immune system destroys the cells in the pancreas that produce insulin, a hormone utilized by the human body to maintain and regulate blood glucose levels. Type 2 diabetes, caused by dietary and other environmental factors, is a health issue in which cells become overwhelmed by insulin and fail to properly respond and they ultimately become insulin resistant. �
Long-term diabetes, either type 1 or type 2, can have many consequences for the brain cells, or neurons, as well as the brain. High blood glucose levels can affect the brain�s functional connectivity which connects brain regions that share functional properties and brain matter. It can also cause the brain to atrophy or shrink and it can lead to small-vessel disease, which restricts blood flow in the brain, causing cognitive difficulties and it can cause the development of vascular dementia. �
In her laboratory, Novak evaluated several ways to prevent these effects in people with type 2 diabetes. One of these ways involves a nasal spray known as intranasal insulin (INI). When used, INI enters the brain and binds to receptors in its memory networks, including the hippocampus, hypothalamus, and insular cortex. As signaling within these memory networks become more efficient, cognitive functions in these areas, such as learning and visual perceptions of spatial relationships, improve. �
�Type 2 diabetes accelerates brain aging,� says Novak, �which, in turn, accelerates the progression of functional decline. With intranasal insulin, we�re hoping to find a new avenue for treatment to slow down these effects or prevent them altogether.� �
In a pilot research study, Novak and her colleagues found that a single dose of INI had a positive effect on memory, verbal learning, and spatial orientation. She is now planning the first clinical trial of INI in older adults with type 2 diabetes. The results of the clinical trial are especially relevant because of the high prevalence of dementia and significant cognitive decline among older adults with diabetes. Sugar, or glucose, is fundamental, however, it must be controlled for overall brain health. �
Glucose, or sugar, is an important source of energy fuel for every cell in the human body, especially the brain. However, excess amounts of blood glucose, or sugar, levels can be more harmful than beneficial and it can ultimately cause a variety of brain health issues, including neurological diseases like dementia and Alzheimer’s disease. Dysglycemia, or abnormal blood glucose, or sugar, levels, is a common condition in diabetes. Managing and regulating glucose, or sugar, in patients with diabetes is essential to promote overall brain health and wellness, according to research studies. – Dr. Alex Jimenez D.C., C.C.S.T. Insight
Neurotransmitter Assessment Form
The following Neurotransmitter Assessment Form can be filled out and presented to Dr. Alex Jimenez. Symptoms listed on this form are not intended to be utilized as a diagnosis of any type of disease, condition, or any other type of health issue. �
Do you often feel low brain endurance for focus and concentration? Do you often crave sugar and sweets in the afternoon? Or do you feel energized after meals? Glucose, or sugar, is the main source of energy in the human body. And, because the human brain has so many nerve cells or neurons, it is one of the most energy-demanding organs, which utilizes about one-half of all the energy from glucose in the human body. Sugar is important but too much of it can also have its downsides. �
Brain functions, such as memory, thinking, and learning, are relatively associated with glucose levels and how efficiently the brain utilizes this essential energy fuel source. If there isn�t enough glucose, or sugar, in the brain, by way of instance, neurotransmitters, or the human brain�s chemical messengers, don’t develop properly and the communications between neurons can ultimately break down. Additionally, dysglycemia, a common health issue caused by abnormal blood glucose levels, can cause loss of energy for brain function and has also been associated with poor attention and cognitive function. �
The scope of our information is limited to chiropractic, musculoskeletal, and nervous health issues or functional medicine articles, topics, and discussions. We use functional health protocols to treat injuries or disorders of the musculoskeletal system. Our office has made a reasonable attempt to provide supportive citations and has identified the relevant research study or studies supporting our posts. We also make copies of supporting research studies available to the board and or the public upon request. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900.�
Curated by Dr. Alex Jimenez �
References:
Marchione, Victor. �Cognition and Brain Structure Affected by Dysglycemia in Older Adults: Study.� Bel Marra Health – Breaking Health News and Health Information, Bel Marra Health, 10 Jan. 2017, www.belmarrahealth.com/cognition-brain-structure-affected-dysglycemia-older-adults-study/.
Edwards, Scott. �Sugar and the Brain.� Sugar and the Brain | Department of Neurobiology, neuro.hms.harvard.edu/harvard-mahoney-neuroscience-institute/brain-newsletter/and-brain-series/sugar-and-brain.
Additional Topic Discussion: Chronic Pain
Sudden pain is a natural response of the nervous system which helps to demonstrate possible injury. By way of instance, pain signals travel from an injured region through the nerves and spinal cord to the brain. Pain is generally less severe as the injury heals, however, chronic pain is different than the average type of pain. With chronic pain, the human body will continue sending pain signals to the brain, regardless if the injury has healed. Chronic pain can last for several weeks to even several years. Chronic pain can tremendously affect a patient’s mobility and it can reduce flexibility, strength, and endurance.
Neural Zoomer Plus for Neurological Disease
Dr. Alex Jimenez utilizes a series of tests to help evaluate neurological diseases. The Neural ZoomerTM Plus is an array of neurological autoantibodies which offers specific antibody-to-antigen recognition. The Vibrant Neural ZoomerTM Plus is designed to assess an individual�s reactivity to 48 neurological antigens with connections to a variety of neurologically related diseases. The Vibrant Neural ZoomerTM Plus aims to reduce neurological conditions by empowering patients and physicians with a vital resource for early risk detection and an enhanced focus on personalized primary prevention. �
Food Sensitivity for the IgG & IgA Immune Response
Dr. Alex Jimenez utilizes a series of tests to help evaluate health issues associated with food sensitivities. The Food Sensitivity ZoomerTM is an array of 180 commonly consumed food antigens that offers very specific antibody-to-antigen recognition. This panel measures an individual�s IgG and IgA sensitivity to food antigens. Being able to test IgA antibodies provides additional information to foods that may be causing mucosal damage. Additionally, this test is ideal for patients who might be suffering from delayed reactions to certain foods. Utilizing an antibody-based food sensitivity test can help prioritize the necessary foods to eliminate and create a customized diet plan around the patient�s specific needs. �
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
�
For your convenience and review of the XYMOGEN products please review the following link. *XYMOGEN-Catalog-Download �
* All of the above XYMOGEN policies remain strictly in force.
Do you often feel energy level drops in the afternoon? Do you often crave sugar and sweets in the afternoon? Do you often have difficulty concentrating before eating? Various medical conditions can affect the overall health of our body and mind. However, research studies have found that anemia caused by iron deficiency can tremendously affect our brain health. �
Iron deficiency is considered to be one of the most prevalent nutritional health issues, affecting approximately 2.5 billion people worldwide. In developing countries, about 40 percent of children and 50 percent of pregnant women have an iron deficiency. Iron is an essential mineral found in approximately 5 percent of the earth�s crust, however, inefficiency in absorption, low iron levels in staple grain foods, and a variety of medical conditions can make iron deficiency a common problem among humans. In first world countries, iron deficiency is still one of the most common nutrient deficiencies. �
What Causes Iron Deficiency and Anemia?
Poor iron intake and increased iron loss, generally through bleeding or breastfeeding, are several of the main causes of iron deficiency. Pregnant women, breastfeeding women, women with heavy periods, children or picky eaters, vegetarians and vegans, as well as people with digestion health issues which cause decreased iron absorption like celiac disease or post gastric bypass, and people with increased bleeding, such as cancer, ulcers, gastritis, or parasites, are generally at higher risk for iron deficiency. High calcium intake, by way of instance, children who drink a lot of milk, can also affect iron absorption, together with drugs and/or medications, such as antacids and proton-pump inhibitors for gastroesophageal reflux disease. �
Although low iron levels are well-known for causing anemia because red blood cells need iron as a part of hemoglobin, iron is also needed for the brain and nerves. Severe iron deficiency in younger children can ultimately cause irreversible damage to cognition and result in lower IQ and developmental delays, especially during the most fundamental stages of human development and up to 16 months of age. Even in adults. the most common symptoms associated with iron deficiency are generally neurological symptoms, including fatigue, brain fog, and restless legs that cause insomnia, among other symptoms. �
Pica, the abnormal behavioral compulsion to eat non-nutritional foods like dirt or clay, is tremendously prevalent in regions of the world where iron deficiency is common. In the developed world, pica is a rare health issue, however, it still frequently occurs in children, pregnant women, and among other groups of people that are at higher risk for iron deficiency, including people who have had gastric bypass. Non-neurological symptoms associated with iron deficiency ultimately include pallor, generalized weakness, and higher than usual heart rate along with shortness of breath, especially with exertion. �
What are the Symptoms of Iron Deficiency?
As previously mentioned above, iron deficiency can cause problems associated with cognition and neurological health issues, such as restless legs and insomnia. The exact mechanisms of why this happens are unknown, however, without enough iron in the brain and nerves, there are problems with neurotransmitter signaling, the development of nerve insulation known as myelin, and brain energy metabolism. Reduced central neuron processing is considered to be one of the most critical problems associated with iron deficiency, which can be a cause of psychiatric symptoms and ongoing psychiatric problems. �
Occasionally, iron deficiency may also cause anxiety, depression, irritability, and even poor concentration and restlessness. By way of instance, iron deficiency has a much higher prevalence in children with ADHD but the symptoms can improve with iron supplements. People with iron deficiency have higher risks of developing psychiatric disorders, especially ADHD, and developmental disorders. Evidence has demonstrated that iron deficiency can cause a variety of other health issues. �
Iron enters the brain through the blood-brain barrier via transferrin receptors. Iron uptake into the brain is highly regulated but it also does highly depend on the iron status of the human body. Therefore, people with low iron levels will have much less iron going into the brain and people with high iron levels will have much more iron going into the brain. Several regions of the brain also appear to gather iron and have higher levels than others. Moreover, neurological symptoms can manifest before developing iron deficiency anemia. Thus, healthcare professionals can’t rule out iron deficiency anemia from the most commonly utilized basic screening test, a complete blood count. A better general screen involves ferritin levels, where less than 15 ng/ml presents the diagnosis for iron deficiency but less than 40 ng/ml presents with fatigue, brain fog, restless legs, and other neurological symptoms. Ferritin on its own can be misleading in populations of people with chronic inflammation, including people on dialysis, where ferritin can be high even if the person is diagnosed with iron deficiency. Furthermore, a full iron workup includes hemoglobin, MCV, ferritin, total iron-binding capacity, serum iron, and transferrin saturation. �
What is the Treatment for Iron Deficiency Anemia?
Treating iron deficiency is considerably simple through the utilization of iron supplements or in mild or moderate cases by encouraging the consumption of foods that are high in iron. Occasionally, people with severe absorption health issues will need iron transfusions intravenously. Meat and seafood are the best sources of easily absorbable heme iron, however, non-heme iron is naturally found in leafy greens, beans, and nuts. Make sure to talk to your doctor if you have iron deficiency. �
It is ultimately essential to make sure if you have iron deficiency before treating it with increased amounts of iron supplements. With the exception of blood loss, the only way to reduce excess iron is through the process of skin cells flaking off. Therefore, adult men who take a lot of iron supplements and people with a genetic tendency to absorb more iron from foods are at a higher risk of developing a medical condition, known as hemochromatosis or severe iron overload. �
Excess iron is stored in the liver and can lead to scarring of the liver, known as cirrhosis. Iron overload can also lead to joint and hormonal problems and it can also cause a bronze-ish skin color. Symptoms of hemochromatosis include joint pain, fatigue, and low sex drive as well as a higher risk of developing diabetes. High serum iron is associated with health issues like high blood pressure. People who aren�t iron deficient can consider regular blood donations to prevent accidental iron overload. Iron is one of those types of minerals that should neither be too high or too low but rather, just right. More research studies, especially clinical trials analyzing common medical conditions, such as restless legs, insomnia, and ADHD, are fundamental to help healthcare professionals understand the relationship between iron deficiency anemia and brain health. �
Recent research studies have demonstrated that iron deficiency anemia may be associated with brain health issues. Because the brain and nerves need iron for many functions, iron deficiency can cause a variety of symptoms and medical conditions, including brain fog, fatigue, restless legs with insomnia, anxiety, depression, and cognitive problems, besides anemia or lack of healthy red blood cells. Treatment for iron deficiency anemia may utilize iron supplements, however, it’s important to make sure to talk to a qualified healthcare professional in order to avoid risks and side effects through iron supplementation.� – Dr. Alex Jimenez D.C., C.C.S.T. Insight
Neurotransmitter Assessment Form
The following Neurotransmitter Assessment Form can be filled out and presented to Dr. Alex Jimenez. Symptoms listed on this form are not intended to be utilized as a diagnosis of any type of disease, condition, or any other type of health issue. �
Do you often feel energy level drops in the afternoon? Do you often crave sugar and sweets in the afternoon? Do you often have difficulty concentrating before eating? Various medical conditions can affect the overall health of our body and mind. However, research studies have found that anemia caused by iron deficiency can tremendously affect our brain health. � Iron deficiency is considered to be one of the most prevalent nutritional health issues, affecting approximately 2.5 billion people worldwide. In developing countries, about 40 percent of children and 50 percent of pregnant women have an iron deficiency. Iron is an essential mineral found in approximately 5 percent of the earth�s crust, however, inefficiency in absorption, low iron levels in staple grain foods, and a variety of medical conditions can make iron deficiency a common problem among humans. In first world countries, iron is still considered to be the most common nutrient deficiency. �
The scope of our information is limited to chiropractic, musculoskeletal, and nervous health issues or functional medicine articles, topics, and discussions. We use functional health protocols to treat injuries or disorders of the musculoskeletal system. Our office has made a reasonable attempt to provide supportive citations and has identified the relevant research study or studies supporting our posts. We also make copies of supporting research studies available to the board and or the public upon request. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900.�
Curated by Dr. Alex Jimenez �
References:
Deans, Emily. �Heavy Metal: Iron and the Brain.� Psychology Today, Sussex Publishers, 29 Nov. 2015, www.psychologytoday.com/us/blog/evolutionary-psychiatry/201511/heavy-metal-iron-and-the-brain.
Additional Topic Discussion: Chronic Pain
Sudden pain is a natural response of the nervous system which helps to demonstrate possible injury. By way of instance, pain signals travel from an injured region through the nerves and spinal cord to the brain. Pain is generally less severe as the injury heals, however, chronic pain is different than the average type of pain. With chronic pain, the human body will continue sending pain signals to the brain, regardless if the injury has healed. Chronic pain can last for several weeks to even several years. Chronic pain can tremendously affect a patient’s mobility and it can reduce flexibility, strength, and endurance.
Neural Zoomer Plus for Neurological Disease
Dr. Alex Jimenez utilizes a series of tests to help evaluate neurological diseases. The Neural ZoomerTM Plus is an array of neurological autoantibodies which offers specific antibody-to-antigen recognition. The Vibrant Neural ZoomerTM Plus is designed to assess an individual�s reactivity to 48 neurological antigens with connections to a variety of neurologically related diseases. The Vibrant Neural ZoomerTM Plus aims to reduce neurological conditions by empowering patients and physicians with a vital resource for early risk detection and an enhanced focus on personalized primary prevention. �
Food Sensitivity for the IgG & IgA Immune Response
Dr. Alex Jimenez utilizes a series of tests to help evaluate health issues associated with food sensitivities. The Food Sensitivity ZoomerTM is an array of 180 commonly consumed food antigens that offers very specific antibody-to-antigen recognition. This panel measures an individual�s IgG and IgA sensitivity to food antigens. Being able to test IgA antibodies provides additional information to foods that may be causing mucosal damage. Additionally, this test is ideal for patients who might be suffering from delayed reactions to certain foods. Utilizing an antibody-based food sensitivity test can help prioritize the necessary foods to eliminate and create a customized diet plan around the patient�s specific needs. �
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
�
For your convenience and review of the XYMOGEN products please review the following link. *XYMOGEN-Catalog-Download �
* All of the above XYMOGEN policies remain strictly in force.
How often do you feel agitated, easily upset, and nervous between meals? How often do you depend on coffee to keep yourself going?How often do you have difficulty concentrating before eating? Inflammation is an essential reaction of the human body. It’s triggered by the immune system to protect us from injury, infection, and/or illness. However, what happens if there is too much inflammation in the human body? And, what happens if there is too much inflammation in the brain?
Neuroinflammation can cause a variety of health issues, such as anxiety, stress, depression, brain fog, fatigue, and even lethargy, among other well-known symptoms. Fortunately, there is one natural remedy that can help tremendously reduce inflammation and improve brain function. According to research studies, curcumin can help combat neuroinflammation. The purpose of the article below is to discuss the anti-inflammatory effects of curcumin in microglia, brain health, and wellness.
Anti-inflammatory Effects of Curcumin in Microglial Cells
Abstract
Lipoteichoic acid (LTA) induces neuroinflammatory molecules, contributing to the pathogenesis of neurodegenerative diseases. Therefore, suppression of neuroinflammatory molecules could be developed as a therapeutic method. Although previous data supports an immune-modulating effect of curcumin, the underlying signaling pathways are largely unidentified. Here, we investigated curcumin�s anti-neuroinflammatory properties in LTA-stimulated BV-2 microglial cells. Inflammatory cytokine tumor necrosis factor-? [TNF-?, prostaglandin E2 (PGE2), and Nitric Oxide (NO] secretion in LTA-induced microglial cells were inhibited by curcumin. Curcumin also inhibited LTA-induced inducible NO synthases (iNOS) and cyclooxygenase-2 (COX-2) expression. Subsequently, our mechanistic studies revealed that curcumin inhibited LTA-induced phosphorylation of mitogen-activated protein kinase (MAPK) including ERK, p38, Akt, and translocation of NF-?B. Furthermore, curcumin induced hemeoxygenase (HO)-1HO-1 and nuclear factor erythroid 2-related factor 2 (Nrf-2) expression in microglial cells. Inhibition of HO-1 reversed the inhibition effect of HO-1 on inflammatory mediators released in LTA-stimulated microglial cells. Taken together, our results suggest that curcumin could be a potential therapeutic agent for the treatment of neurodegenerative disorders via suppressing neuroinflammatory responses. � Keywords:curcumin, neuroinflammation, TLR2, HO-1, microglial cells
Introduction
Chronic neuroinflammation plays an important role in various neurodegenerative diseases, including AD, Parkinson�s disease (PD), Huntington�s disease (HD), stroke, amyotrophic lateral sclerosis (ALS), and multiple sclerosis (MS) (Spangenberg and Green, 2017). Neuroinflammation is interceded by the activation of microglia, the prime effector cells and resident immune cells of the CNS (Nakagawa and Chiba, 2015). Microglial cells can be activated in response to neuronal death or neuronal damage induced by neuroinflammatory responses or by extracellular toxins, such as bacteria and pathogens (Larochelle et al., 2015). In neuroinflammation, activated microglia releases various kinds of cytokines, chemokines, reactive oxygen species, and reactive nitrogen species for the development and maintenance of inflammatory responses (Moss and Bates, 2001). Excessive production of these inflammatory mediators could cause neuronal damage and death. Accumulated evidence suggests that control of microglial activation could attenuate the severity of neurodegenerative disease (Perry et al., 2010). Therefore, the development of anti-neuro-inflammatory agents for the inhibition of microglial activation could be beneficial for the treatment of neurodegenerative diseases.
Microglia express pattern recognition receptors (PRR) that can bind to pattern-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) such as lipopolysaccharide (LPS) and lipoteichoic acid (LTA), respectively (Jack et al., 2005). TLRs, a major class of PRRs, play a crucial role in host defense by inducing innate immune responses. Increasingly, studies have indicated that TLR2 agonist LTA is involved in the pathogenesis of CNS infectious diseases and can induce neuronal damage (Neher et al., 2011). Inhibition of TLR2 activation attenuates microglial cell activation and amyloid ? accumulation in the brain (McDonald et al., 2016; Hossain et al., 2017). Signal transduction via TLR2 is mediated by different adaptor proteins, including MyD88, which promotes downstream signaling via MAPK and NF-?B activation leading to the expression of inflammatory mediators (Larochelle et al., 2015).
Inflammatory and oxidative molecules are very potent activators of Keap-Nrf2 (NF-E2-related factor 2), which induces the expression of Phase II detoxification enzymes to adapt to the oxidative stress condition (Rojo et al., 2010). Usually, Nrf2 acts in an inactive form. Upon stimulation, Nrf2 separates from Keap1 and translocates into the nucleus, where it binds to the antioxidant response element (ARE) to activate the transcription of antioxidant genes for cytoprotection (Ma, 2013; Cho et al., 2015). One of the Nrf2-regulated genes is heme oxygenase-1 (HO-1), which has an ARE sequence in its promoter region. Recently, HO-1 has been reported to be a predominant factor in controlling oxidative stress and inflammatory responses in neurodegenerative diseases (Schipper et al., 2009). HO-1 is the first inducible rate-limiting enzyme in the degradation of heme into by-products. HO-1 may provide neuroprotection or neurotoxic effect because of the balance between the beneficial and toxic effects of heme and heme products (Mancuso et al., 2010). One by-product of HO-1, Bilirubin, has been demonstrated to protect neurons from oxidative stress in vivo and in vitro. Bilirubin can be oxidized to biliverdin by scavenging peroxyl radicals (Chen, 2014). It has been suggested that HO-1, biliverdin, and CO have anti-inflammatory properties (Jazwa and Cuadrado, 2010). Another study has suggested that mice lacking HO-1 were vulnerable to pro-inflammatory stimuli and developed chronic inflammation due to reduced iron levels (Chora et al., 2007). Furthermore, a recent study suggested that the up-regulation of the Nrf2 and HO-1 pathways significantly inhibited the inflammatory reaction in activated microglia (Kim et al., 2016). Nrf2 inhibited microglial hyperactivation by suppressing p38 MAPK and the NF-?B signaling pathway (Kim B.W. et al., 2013). Knockdown of Nrf2 in mice was shown to be hypersensitive to neuroinflammation, as indicated by an increase in the inflammatory markers iNOS, IL-6, and TNF-? (Rojo et al., 2010). Consequently, Nrf2 and HO-1 have been considered as important therapeutic targets for neurodegenerative diseases (Koh et al., 2011; Zhang et al., 2014).
Curcumin, the main curcuminoid isolated from Curcuma longa L. (turmeric) has been used for centuries in Southeast Asia both as a medicinal remedy and as food (Kunnumakkara et al., 2017). Curcumin, demethoxycurcumin, bisdemethoxycurcumin, ar-turmerone, ?-turmerone, and ?-turmerone are the major bioactive compounds found in C. longa. In modern pharmacological studies, C. longa constituents, particularly curcumin, have shown promising pharmacological activities due to its anti-neuroinflammatory, neuroprotective, chemopreventive, immunomodulatory, and potentially chemotherapeutic effects (Garcia-Alloza et al., 2007; Zhou et al., 2017). A previous study showed that curcumin inhibited LPS-induced inflammatory responses in RAW264.7 macrophages, suggesting a potential role of curcumin in anti-Gram-negative bacterial infection (Zhou et al., 2017) and both in vivo and in vitro research have shown that curcumin exhibits anti-inflammatory effects (Garcia-Alloza et al., 2007; Prakobwong et al., 2011; Parada et al., 2015; Li et al., 2016). Furthermore, curcumin has also been reported to promote the development of the M2 microglial phenotype in an HO-1-dependent manner and reduce iNOS induction, protecting microglial cells against oxidative stress (Parada et al., 2015). In the present study, we investigated whether curcumin could affect LTA-induced microglial activation. The TLR2 ligand LTA is a major constituent of the cell wall of Gram-positive bacteria. We show that curcumin exhibits anti-inflammatory and antioxidant effects in LTA-stimulated BV2 microglia through activation of HO-1/Nrf2/ARE cytoprotective mechanisms.
Materials and Methods
Materials
Curcumin and other reagents were purchased from Sigma (C7727, >80%, St. Louis, MO, United States). Protoporphyrin IX (SnPP) and antibodies directed against HO-1 (sc-390991) – Nrf2 (sc-722), TATA-binding protein (TBP; sc-74595), ?-tubulin (sc-134237), and ?-actin (sc-130065) – were purchased from Santa Cruz Biotechnology, Inc., (Dallas, TX, United States). Antibodies directed against iNOS (13120) – phosphorylated (p)-MAPK (9910s), MAPK (9926), protein kinase B (Akt; 4685), p-Akt (13038), and an NF-?B pathway kit (9936) – were purchased from Cell Signaling Technology, Inc., (Danvers, MA, United States). LTA was obtained from InvivoGen (tlrl-pslta,Toulouse, France). Additionally, JNK inhibitor (JNK inhibitor II; 420119), Akt inhibitor (wortmannin; 12-338), ERK inhibitor (PD98059, 513000), and p38 inhibitor (SB230580, 559395) were purchased from EMD Millipore (Billerica, MA, United States). The cell culture medium, DMEM, and fetal bovine serum (FBS) were purchased from Gibco BRL (now Invitrogen Corporation, Carlsbad, CA, United States).
Cell Culture
Mouse BV-2 microglial cells were purchased from ATCC. Cells were cultured in DMEM supplemented with 10% heat-inactivated FBS and 0.1% penicillin-streptomycin (BioSource International, Camarillo, CA, United States) at 37�C in a humidified atmosphere of 5% CO2 and 95% air.
Cell Viability Assay
The cytotoxicity of curcumin was assessed using a microculture [3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] (MTT)-based colorimetric assay. Cells were incubated in 24-well plates at a density of 5 � 105 cells per well. The MTT solution (5 ml of 5 mg/ml) was added to each well (final concentration 62.5 mg/ml). After incubation for 3 h at 37�C in 5% CO2, the supernatant was removed and the formazan crystals produced in viable cells were solubilized with 150 ml of dimethylsulfoxide (DMSO). The absorbance of each well was then read at 570 nm using a microplate reader (Wallac 1420; PerkinElmer, Inc., Boston, MA, United States).
Measurement of Nitrite Concentration
NO synthesis in cell cultures was measured by the Griess method with microplate. To measure nitrite, 100-?l aliquots were removed from the conditioned medium and incubated with an equal volume of the Griess reagent [1% sulfanilamide/0.1%N-(1-naphthyl)-ethylenediaminedihydrochloride/2.5% H3PO4] at room temperature for 10 min. The nitrite concentration was determined by measuring the absorbance at 540 nm with a Vmax 96-well microplate spectrophotometer (Molecular Devices, Menlo Park, CA, United States). Sodium nitrite was used as a standard.
Measurement of TNF-? and PGE2 Concentration
The cells were incubated first with various concentrations of curcumin for 1 h and then with LTA for 16 h. Following 24 h incubation, TNF-? and PGE2 levels were quantified in the culture media using an enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN, United States) according to the manufacturer�s instructions.
Preparation of Nuclear Extract
BV-2 microglial cells were washed three times with cold PBS and collected in 3000 ?l PBS using centrifugation at 800 �g for 5 min (4�C). The cell pellets were suspended in buffer A [10 mM HEPES-KOH (pH 7.9); 1.5 mM MgCl2; 10 mM KCl; 0.5 mM dithiothreitol (DTT); 0.2 mM protease inhibitor (PI)] and incubated for 5 min on ice. Buffer B [10 mM HEPES-KOH (pH 7.9); 1.5 mM MgCl2; 420 mM NaCl; 0.2 mM EDTA; glycerol 25% v/v; 0.1 mM DTT; 0.2 mM PI] was added to the cell extract and was incubated on ice for 5 min prior to centrifugation at 11,000 �g for 1 min at 4�C. Nuclear proteins were extracted with the addition of complete lysis buffer B [10 mM HEPES-KOH (pH 7.9); 1.5 mM MgCl2; 10 mM KCl; 0.5 mM DTT; 0.2 mM PI; 25% (w/v) glycerin; 420 mM NaCl; 0.2 mM EDTA] for 30 min at 4�C with occasional vortexing. Following centrifugation at 11,000 �g for 5 min at 4�C, the supernatants were collected and stored at -70�C.
Western Blot Analysis
BV-2 cells were harvested in an ice-cold lysis buffer (1% Triton X-100; 1% deoxycholate; 0.1% sodium dodecyl sulfate). The protein content of the cell lysates was subsequently determined using Bradford reagent (Bio-Rad Protein Assay Kit I5000001; Bio-Rad Laboratories, Inc., Hercules, CA, United States). Total proteins in each sample (50 ?g) were separated by 7.5% SDS-PAGE and transferred to polyvinylidene difluoride membranes. Following blocking of the non-specific binding sites with 5% non-fat milk at room temperature for 30 min, the membranes were incubated with primary antibodies directed against iNOS (1:500), p-Akt (1:1,000), p-MAPK (1:1,000), MAPK (1:1,000), p-p65, p65 (1:500), p-I?B?, I?B? (1:1,000), HO-1 (1:1,000), Nrf2 (1:1,000), TBP (1:3,000), ? (1:1,000), HO-1 (1:1.0), and actin (1:3,000) for 16 h at 4�C. This was followed by incubation with horseradish peroxidase-conjugated anti-rabbit (sc-2768; 1:5,000) or anti-mouse (sc-2371; 1:5,000) secondary antibodies (Santa Cruz Biotechnology, Inc.) at room temperature for 1 h. Tubulin was used as the loading control for each lane. The proteins were visualized using an enhanced chemiluminescence detection kit (GE Healthcare, Chicago, IL, United States). Following washing with PBS with Tween-20, the protein bands were visualized using the Gel Docsed as the loading control for each lane. The proteins were visualized using a Quant 350 analyzer (GE Healthcare).
Real-Time RT-PCR
Total RNA was isolated from cells using an RNA spin miniRNA isolation kit (GE Healthcare, Uppsala, Sweden) according to the manufacturer�s instructions. cDNA was synthesized from 1 ?g of total RNA using Maxime RT PreMix (Takara, Gyeonggi-do, Japan) and anchored oligo-dT15-primers. Real-time PCR was performed using a Chromo4TM instrument (Bio-Rad) and SYBR Green Master Mix (Applied Biosystems, Foster City, CA, United States). Relative amounts of target mRNA were determined using the comparative threshold (Ct) method by normalizing target mRNA Ct values to those for ?-actin (Ct). Prime sequences used in the study were shown in Table ?1.
Statistical Analysis
Data are expressed as the mean (standard deviation, SD). Each experiment was repeated at least three times. Statistical analysis was performed using the Statistical Package for GraphPad Prism software (version 16.0) to determine significant differences. We used either Student�s t-test or one-way analysis of variance (ANOVA) followed by Dunn�s post hoc tests for analyses. P-values < 0.05 were considered statistically significant.
Results
Curcumin Did Not Affect Cell Viability
Cell viability experiments were carried out to determine whether concentrations of curcumin used in this study affected the viability of BV2 microglia. Figure ?1 shows that curcumin at the concentration range of 5�20 ?M, together with or without 5 ?g/ml LTA, did not produce cytotoxicity in BV2 microglia. Therefore, we used these concentrations of curcumin for further study.
Curcumin Prevented the Production of Neuroinflammatory Molecules in LTA-Activated BV2 Microglia
To investigate the effects of curcumin on the secretion of inflammatory cytokines, BV2 cells were treated with LTA in the presence and absence of curcumin for 24 h. Curcumin was not removed before LTA addition. Release of NO, PGE2, and TNF-? were significantly and dose-dependently reduced by curcumin (Figures 2A�C). Furthermore, LTA increased the mRNA expression of iNOS and COX-2. Incubation with curcumin suppressed the mRNA expression of COX-2 and iNOS in BV2 microglial cells stimulated by LTA in a concentration-dependent manner (Figures 2D, E).
Curcumin Suppressed LTA-Induced Activation of NF-?B in BV-2 Microglial Cells
The genes encoding inflammatory protein expression in response to microglial activation were under the transcription control of NF-?B. Therefore, we examined the effect of curcumin on the activation of NF-?B in LTA-stimulated microglial cells. The results showed that LTA induced a characteristic increase in the phosphorylation of I?B?. Following pre-treatment with curcumin, levels of p-I?B? were significantly reduced in a concentration-dependent manner (Figure ?3 and Supplementary Figure S1). Consistently, the nuclear translocation of the NF-?B p65 subunit induced by LTA was also attenuated by pre-treatment with curcumin. Taken together, curcumin likely attenuates the expression of neuroinflammatory molecules by suppressing the nuclear translocation and activation of NF-?B. Quantification with statistical analysis was provided as supporting data.
Curcumin Inhibited LTA-Induced Activation of p38, and ERK MAPK in BV-2 Microglial Cells
Apart from NF-?B, MAPKs are also upstream modulators of neuroinflammatory molecules in microglial cells. Previous studies showed that curcumin antagonized LPS-induced MAPKs phosphorylation in microphage (Yang et al., 2008; Kunnumakkara et al., 2017). To investigate whether curcumin inhibits neuroinflammation through regulating MAPKs, we examined its effects on LTA-induced MAPK phosphorylation. BV-2 microglial cells were pre-treated with different concentrations of curcumin for 3 h and were then stimulated with LTA for 1 h. As shown in Figure ?4A and Supplementary Figure S2, curcumin inhibited LTA-induced ERK, p38, and Akt phosphorylation. However, up to 20 ?M curcumin did not affect LTA-induced JNK phosphorylation. MAPKs pathway has been reported to mediate the production of cytokines, chemokine, and other neuroinflammatory molecules. Therefore, we next investigated the role of ERK, p38, JNK, and Akt in BV2 cells� neuroinflammatory molecule production using the ERK, p38, JNK, and Akt inhibitors. However, only the p38 inhibitor SB203580 significantly decreased LTA-induced release of NO and mRNA expression levels of iNOS (Figures 4B, C). Although phosphorylation of JNK was not inhibited by curcumin, the JNK inhibitor II significantly inhibited LTA-induced NO release (Figure ?4B). The results suggest that MAPKs� signaling pathways are involved in curcumin�s anti-neuroinflammatory effects in LTA-stimulated microglial. Quantification with statistical analysis is provided as supporting data.
Inhibition of HO-1 Signaling Abolished Curcumin�s Inhibitory Effect on Neuroinflammatory Responses
HO-1 acts as an anti-inflammatory and antioxidant modulator in microglia (Schipper et al., 2009). Western blot and RT-PCR analyses showed that curcumin upregulated HO-1 expression at the protein and mRNA levels, as shown in Figures 5A�D and Supplementary Figure S3. The expression of HO-1 mRNA and protein was maximally increased in BV-2 microglial cells treated with 20?M curcumin for 4 h and 8 h respectively. Furthermore, curcumin increased Nrf2 nuclear translocation within 1 h and prolonged its nuclear translocation state to 2 h (Figures 5E, F and Supplementary Figure S3). Next, we investigated whether curcumin-induced HO-1 mediated an anti-neuroinflammatory response in LTA-stimulated BV-2 microglial cells. We treated cells with the HO-1 inhibitor SnPP. We then evaluated curcumin�s effect on LTA-induced NO and TNF-? release. Treatment with SnPP significantly suppressed curcumin-mediated inhibition of NO and TNF-a release (Figures 5G, H). Taken together, these results reveal that curcumin-dependent HO-1 and Nrf-2 signal activation plays a crucial role in downregulating neuroinflammatory responses. Quantification with statistical analysis is provided as supporting data.
Discussion
Microglia, the major resident macrophages of the CNS, has been reported to be the main effector cells in mediating neuroinflammation and selective neuronal death (Perry et al., 2010). Microglial cells increase the production of neuroinflammatory molecules after exposure to activators such as LPS and LTA via their surface receptors, TLR4 and TLR2, respectively (Perry and Holmes, 2014; Hossain et al., 2017). Increased expression and activation of TLR2 is associated with the progression of neurodegenerative diseases, such as PD and dementia (Dzamko et al., 2017). For example, activation of TLR2 could upregulate ?-synuclein in PD brains and play important roles in the pathogenesis of PD brains (Roodveldt et al., 2013). In addition, Kim C. et al. (2013) also showed that neurodegeneration was attenuated by either knockout or knockdown of TLR2 in rodent PD models. Thus, controlling TLR2-mediated microglia activation and neurotoxicity has been suggested as an important therapeutic approach to treating neurodegenerative diseases. A potential agent in this process could be curcumin, which has been shown to exert neuro-protective and anti-inflammatory effects in various experiment models (Parada et al., 2015; Li et al., 2016). Curcumin is a highly lipophilic natural compound. A previous study has well demonstrated that curcumin is able to cross the blood�brain barrier and that it is mainly concentrated in the hippocampus in the brain (Tsai et al., 2011). Some studies reported that curcumin inhibited HIV-1 gp120-induced neuronal damage and provided anti-neuroinflammatory effects in LPS-induced microglia (Gong et al., 2012). This protective effect of curcumin seems to be dependent on its anti-inflammatory actions. Curcumin could protect neurons against microglia-mediated neurotoxicity while becoming inefficient under microglia-depleted conditions (Park et al., 2001; Yang et al., 2008; Parada et al., 2015). Similar studies in peripheral cells also showed the anti-inflammatory effects of curcumin. Using RAW 264.7 murine macrophages, studies have shown that curcumin inhibited PGE2, NO, and TNF-? release following LPS stimulation (Pae et al., 2008). However, the effects of curcumin on TLR2-induced neuroinflammation in microglial cells are not fully understood.
Regulation of the signaling pathways in activated microglia is important in maintaining CNS homeostasis because deregulated neuroinflammatory responses can result in the death of adjacent neurons through the release of inflammatory molecules, such as cytokines, chemokines, NO, and ROS (Perry and Holmes, 2014; Spangenberg and Green, 2017). For example, excessive NO synthesis under endotoxins results in the formation of reactive nitrogen species and neuronal cell death (Perry et al., 2010). PGE2 has also been shown to contribute to neuronal death through activation of the MAPK/ERK pathway in microglia (Xia et al., 2015). In this present study, we showed that curcumin inhibited the secretion of inflammatory mediators TNF-?, NO, and PGE2, and expression of iNOS and COX-2 in BV2 microglia stimulated with LTA. We further showed that curcumin attenuated these effects of LTA without altering cell survival, suggesting that curcumin is safe and could be considered as a potential therapeutic agent in neuroinflammation.
NF-?B is the main transcription factor which plays critical roles in regulating redox homeostasis. NF-?B is considered the master regulator of microglial inflammatory responses to neuronal injury (Acharyya et al., 2007). Recent studies showed that NF-?B activation controlled the expression of inflammatory molecules, such as NO, PGE2, and TNF-?, and IL-1b production (Acharyya et al., 2007). Therefore, modulation of NF-?B activation is considered a critical way to control microglial activation. The activation of the NF-?B signaling pathway is mediated by the I?B protein. The phosphorylation of I?B results in NF-?B dissociation, which leads to the induction of inflammatory mediators. In this study, it was shown that curcumin produced dual inhibition of phosphorylation and degradation of I?B?, as well as nuclear translocation of p65, suggesting that this agent could stabilize NF-?B in the microglial cytoplasm following stimulation with LTA in BV-2 microglial cells.
In mammalian cells, MAPKs signaling pathways, including ERK, JNK, and p38, contribute to the production of a wide variety of neuroinflammatory mediators (Chantong et al., 2014). In this present study, pre-treatment with curcumin decreased the phosphorylation of p38 and ERK. Furthermore, the p38 inhibitor SB203580 significantly reduced the secretion of NO and the mRNA expression of the key pro-inflammatory gene, iNOS. These results suggested that curcumin initiated the anti-neuroinflammatory effects in LTA-stimulated BV-2 microglial cells, partially through inhibition of p38 MAPK activation. The PI3K/Akt-dependent signaling pathway promotes inflammatory responses in microglia. The involvement of the Akt pathway has been shown in the expression of inflammatory mediators in microglia through the activation of NF-?B in microglia (Lo et al., 2015). Curcumin suppressed the phosphorylated Akt, the downstream target of PI3K. However, the PI3K inhibitor wortmannin did not show any inhibitory effect on the secretion of NO or the mRNA expression of iNOS. Taken together, these data suggest that the anti-neuroinflammatory effect of curcumin occurs mainly through inhibiting the NF-?B and MAPKs signaling.
We also identified the intracellular pathway that negatively regulates the inflammatory-molecule expression in microglial cells. Nrf2 is a redox-sensitive transcription factor that regulates microglial inflammatory responses to brain infections. The effect of Nrf2 has been described in different in vivo models where knockdown of Nrf2 in mice enhanced vulnerability to asthma or emphysema (Ma, 2013). Moreover, the TLR2/TLR4 agonist promoted inflammatory responses in Nrf2 KO mice compared to WT mice (Kong et al., 2011). In the current study, we showed that curcumin increased the expression of Nrf2 and its downstream protein HO-1. HO-1 is a key signaling molecule implicated in the regulation of inflammatory and oxidative responses. The HO-1 gene has an ARE sequence in its promoter region, which is a binding site for the transcription factor Nrf2. Several studies have proposed that NF-?B interrupts the Nrf-2-ARE signaling pathway because many compounds that induced HO-1 and Nrf2 signaling incidentally repressed NF-?B activation (Li et al., 2016). HO-1 expression was essential for the microglial specific cytoprotective effect (Parada et al., 2015). Several studies have also shown an inverse correlation between HO-1 and inflammatory mediator secretion (Chora et al., 2007; Parada et al., 2015). In agreement, we observed that curcumin alone induced the expression of HO-1 in microglial cells. Furthermore, the HO-1 inhibitor abrogated curcumin anti-inflammatory effect in BV-2 microglial cells.
Conclusion
This study demonstrated that curcumin had anti-inflammatory activity in LTA-stimulated microglial cells that may through inhibiting NF-?B and p38 MAPK activation, and may induce the expression of Nrf2 and HO-1 (Figure ?6). Furthermore, curcumin does not have cytotoxic effects in BV-2 microglial cells at its anti-inflammatory dose. Curcumin may have therapeutic potential for some neuroinflammation-associated disorders caused by Gram-positive bacteria.
�
Curcumin, or turmeric, is a powerful anti-inflammatory which has been demonstrated to have many health benefits. Regarded as an antioxidant with anti-cancer, antidepressant, and anti-aging properties, curcumin can do much more than heal wounds and enhance memory. According to research studies, curcumin or turmeric can help reduce neuroinflammation or brain inflammation. This powerful anti-inflammatory can block the production of proinflammatory cytokines and promote overall well-being. – Dr. Alex Jimenez D.C., C.C.S.T. Insight
Neurotransmitter Assessment Form
[wp-embedder-pack width=”100%” height=”1050px” download=”all” download-text=”” attachment_id=”72741″ /]
The following Neurotransmitter Assessment Form can be filled out and presented to Dr. Alex Jimenez. Symptoms listed on this form are not intended to be utilized as a diagnosis of any type of disease, condition, or any other type of health issue.
In honor of Governor Abbott’s proclamation, October is Chiropractic Health Month. Learn more about the proposal.
How often do you feel agitated, easily upset, and nervous between meals? How often do you depend on coffee to keep yourself going?How often do you have difficulty concentrating before eating? Inflammation is an important response of the human body. It’s activated by the immune system to guard us against injury, infection, and/or illness. However, what happens if there is too much inflammation in the human body? And, what happens if there is too much inflammation in the brain?�
Brain inflammation can cause a variety of health issues, such as anxiety, stress, depression, brain fog, fatigue, and even lethargy, among other common symptoms. Fortunately, there is one natural remedy that can help greatly reduce neuroinflammation and improve brain function. According to research studies, curcumin can combat brain inflammation. The purpose of the article above was to discuss the anti-inflammatory effects of curcumin in microglia and brain well-being
The following article has been referenced from the National Center for Biotechnology Information (NCBI). The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues or functional medicine articles, topics, and discussions. We use functional health protocols to treat injuries or disorders of the musculoskeletal system. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 .
Curated by Dr. Alex Jimenez
Additional Topic Discussion: Chronic Pain
Sudden pain is a natural response of the nervous system which helps to demonstrate possible injury. By way of instance, pain signals travel from an injured region through the nerves and spinal cord to the brain. Pain is generally less severe as the injury heals, however, chronic pain is different than the average type of pain. With chronic pain, the human body will continue sending pain signals to the brain, regardless if the injury has healed. Chronic pain can last for several weeks to even several years. Chronic pain can tremendously affect a patient’s mobility and it can reduce flexibility, strength, and endurance.
Neural Zoomer Plus for Neurological Disease
Dr. Alex Jimenez utilizes a series of tests to help evaluate neurological diseases. The Neural ZoomerTM Plus is an array of neurological autoantibodies which offers specific antibody-to-antigen recognition. The Vibrant Neural ZoomerTM Plus is designed to assess an individual�s reactivity to 48 neurological antigens with connections to a variety of neurologically related diseases. The Vibrant Neural ZoomerTM Plus aims to reduce neurological conditions by empowering patients and physicians with a vital resource for early risk detection and an enhanced focus on personalized primary prevention.
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
For your convenience and review of the XYMOGEN products please review the following link. *XYMOGEN-Catalog-Download* All of the above XYMOGEN policies remain strictly in force.
L-glutamate is one of the main excitatory neurotransmitters in the human brain and it plays an essential role in practically all activities of the nervous system. In the following article, we will discuss the general principles of L-glutamate signaling in the brain. Then, we will demonstrate this scheme by describing the different pools of extracellular glutamate, including the synaptic, the perisynaptic, and the extrasynaptic, resulting from vesicular and non-vesicular sources or abnormally located glutamate receptors outside of synapses as well as discuss their possible physiological functions in the human brain. �
Glutamate Signaling in the Brain
According to research studies, the human brain has about a 6 to 7 ?mol/g wet weight of L-glutamate. L-glutamate, together with glutamine, is one of the most abundant free amino acids in the central nervous system (CNS). More than five decades ago, several research studies demonstrated that L-glutamate has an excitatory response on nerve cells. Since then, its role as an excitatory neurotransmitter as well as its cerebral metabolism has been evaluated in numerous research studies. �
L-glutamate is commonly found throughout synaptic vesicles in the presynaptic terminal through the process of vesicular glutamate transporters. Additionally, several of the L-glutamate in the vesicles may develop by a vesicle-associated aspartate amino-transferase from 2-oxoglutarate utilizing L-aspartate as the amino group donor. During the depolarization of the presynaptic membrane, L-glutamate is released into the synaptic cleft and connects to ionotropic glutamate receptors, known as iGluRs, at the postsynaptic membrane, as shown in Figure 1. According to research studies, iGluRs are characterized as ligand-gated ion channels which include receptors of the ?-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA), kainate, and N-methyl-D-aspartic acid (NMDA) types. While AMPA and kainate receptors primarily regulate and maintain sodium influx, NMDA receptors actually have a high calcium conductivity. Moreover, the activation of NMDA receptors plays a fundamental role in synaptic plasticity and learning. In contrast to the other iGluRs, the activity of NMDA receptors is ultimately restricted by an Mg+2 block at the regular membrane potential, however, the ion channel is immediately unblocked by membrane depolarization which eliminates Mg+2 from the pore. Furthermore, NMDA receptors are tetramers that have two NR1 subunits and two NR2 or NR3 subunits, according to several research studies. �
Additionally to iGluRs, there are also eight isoforms of metabotropic glutamate receptors (mGluRs) which belong to the family of G-protein-coupled receptors, where they don’t develop ion channels but instead signal through a variety of second messenger systems. L-glutamate-associated depolarization causes a postsynaptic excitatory potential which eases the development of an action potential at the axon hillock. The glutamatergic synapse is activated by astrocytic processes that demonstrate high levels of excitatory amino acid transporters (EAATs). There are five different EAATs, EAAT1 to 5, of which EAAT1 and 2 are the primary astrocytic EAATs, whereas EAAT3 shows a predominantly neuronal expression. Approximately 90 percent of the L-glutamate transport is regulated and maintained by EAAT2 such as GLT-1 in rodent models. These transporters then co-transport 2 or 3 molecules of Na+ and a proton with each molecule of L-glutamate or L-aspartate together with the counter-transport of a K+ ion. Therefore, by utilizing the electrochemical gradient of these ions throughout the plasma membrane as an energy source, the transporters are able to safely and effectively accumulate L-glutamate and L-aspartate in cells against their sudden intra- to extracellular concentration gradients. This allows the brain to control a very low extracellular L-glutamate concentration in the low micromolar range. It is generally believed that L-glutamate taken up by astrocytes is turned to glutamine by the enzyme glutamine synthetase, the glutamine is then released, taken up by neurons and turned to L-glutamate, where it is ultimately utilized once again for neurotransmission. �
Extrasynaptic Glutamate in the Brain
Aside from the essential role of L-glutamate as the primary excitatory neurotransmitter released from glutamatergic presynapses, as previously mentioned above, it has become evident that L-glutamate receptors outside the synaptic cleft also play an essential role in brain physiology. In the cerebellum, it was demonstrated by evaluating AMPA receptor-mediated currents in Bergmann glia that synaptically released L-glutamate concentrations can reach extrasynaptic concentrations of up to 190 ?M while concentrations in the synaptic cleft can exceed 1 mM. Moreover, several mGluRs have been shown to demonstrate a different localization in proximity to the postsynaptic density which would allow them to immediately recognize L-glutamate escaping from the synaptic cleft, as shown in Figure 1. However, current research studies have demonstrated that iGluRs, especially of the NMDA type, are also found at extrasynaptic regions in the neuronal cell membrane. Utilizing light and electron microscopy, other research studies also demonstrated that extrasynaptic NMDA receptors gather at different regions of close contact in the dendritic shaft with axons, axon terminals, or astrocytic processes. The proportion of extrasynaptic NMDA receptors was estimated to be as high as 36 percent of the dendritic NMDA receptor pool in rat hippocampal slices. Although extrasynaptic NMDA receptors were associated with similar scaffolding proteins as synaptic NMDA receptors, an in vitro research study suggested that extrasynaptic and synaptic NMDA receptors may ultimately activate different downstream signaling pathways with a variety of results, including the suppression of CREB activity by extrasynaptic NMDA receptor activation as well as activation by synaptic NMDA receptors. Furthermore, NMDA receptors localized extrasynaptically on dendritic shafts connect extrasynaptic L-glutamate as well as regulate and maintain Ca2+ influx during the elimination of the Mg+2 block by dendrite depolarization throughout the backfiring of action potentials. Research studies demonstrated that L-glutamate release from astrocytes can activate slow inward currents through extrasynaptic NMDAR receptors in CA1 neurons which can also be ultimately synchronized. The mechanisms through which glial cells release L-glutamate as well as how the extrasynaptic L-glutamate concentrations are controlled are vital towards understanding how the activity of extrasynaptic NMDA receptors is controlled. �
Different mechanisms through which astrocytes can release L-glutamate have been suggested, including vesicular L-glutamate release and non-vesicular release through anion channels as well as connexin hemichannels and release through the cystine/glutamate antiporter system x?c. Several research studies strongly suggest that vesicular release from astrocytes plays a minor role because the Ca+2-associated release of L-glutamate was still present in astrocytes created from dominant-negative SNARE mice where vesicular release can be blocked by doxycycline withdrawal. System x?c is a cystine/glutamate antiporter which is characterized as heterodimeric amino acid transporters, made up of xCT as the specific subunit and 4F2hc as the promiscuous heavy chain. This transporter is demonstrated in the brain, especially in astroglial and microglial cells, as shown in Figure 1. The fact that extrasynaptic L-glutamate levels in different regions of the human brain are downregulated by approximately 60 percent to 70 percent in xCT knock out mice, research studies demonstrated that system x?c releases L-glutamate into the extrasynaptic space and suggests that this transporter is essential in the regulation of extrasynaptic L-glutamate levels. This is further supported by the observation that when measured by in vivo microdialysis, the increase in extrasynaptic L-glutamate developed by EAAT inhibitors is neutralized by blocking system x?c while blocking neuronal vesicular L-glutamate release is ineffective. Further research studies are still required. �
Taken together, glutamatergic neurotransmissions don’t simply happen through classical excitatory synapses but also through extrasynaptic L-glutamate receptors, as shown in Figure 1. Finally, the levels of extrasynaptic L-glutamate are determined, at least partially, by glial non-vesicular L-glutamate release, as also shown in Figure 1. However, the regulation of extrasynaptic L-glutamate levels, as well as its temporal-spatial dynamics and its effect on neuronal function, neurodegeneration, and behavior, are far from being fully understood by researchers, healthcare professionals, and patients. �
Glutamate, together with aspartate, is one of the main excitatory neurotransmitters in the human brain. Although it plays a fundamental role in the overall structure and function of the nervous system, excessive amounts of glutamate can ultimately cause excitotoxicity which may lead to a variety of health issues, such as Alzheimer’s disease and other types of neurological diseases. The following article describes the role of glutamate in the human brain. – Dr. Alex Jimenez D.C., C.C.S.T. Insight
L-glutamate is one of the main excitatory neurotransmitters in the human brain and it plays an essential role in practically all activities of the nervous system. In the article above, we discussed the general principles of L-glutamate signaling in the brain. Then, we demonstrated this scheme by describing the different pools of extracellular glutamate, including the synaptic, the perisynaptic, and the extrasynaptic, resulting from vesicular and non-vesicular sources or abnormally located glutamate receptors outside of synapses as well as discussed their possible physiological functions in the human brain. The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues as well as functional medicine articles, topics, and discussions. We use functional health protocols to treat injuries or chronic disorders of the musculoskeletal system. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 . �
Curated by Dr. Alex Jimenez �
References �
Lewerenz, Jan, and Pamela Maher. �Chronic Glutamate Toxicity in Neurodegenerative Diseases-What Is the Evidence?� Frontiers in Neuroscience, Frontiers Media S.A., 16 Dec. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4679930/.
Additional Topic Discussion: Chronic Pain
Sudden pain is a natural response of the nervous system which helps to demonstrate possible injury. By way of instance, pain signals travel from an injured region through the nerves and spinal cord to the brain. Pain is generally less severe as the injury heals, however, chronic pain is different than the average type of pain. With chronic pain, the human body will continue sending pain signals to the brain, regardless if the injury has healed. Chronic pain can last for several weeks to even several years. Chronic pain can tremendously affect a patient’s mobility and it can reduce flexibility, strength, and endurance.
Neural Zoomer Plus for Neurological Disease
�
Dr. Alex Jimenez utilizes a series of tests to help evaluate neurological diseases. The Neural ZoomerTM Plus is an array of neurological autoantibodies which offers specific antibody-to-antigen recognition. The Vibrant Neural ZoomerTM Plus is designed to assess an individual�s reactivity to 48 neurological antigens with connections to a variety of neurologically related diseases. The Vibrant Neural ZoomerTM Plus aims to reduce neurological conditions by empowering patients and physicians with a vital resource for early risk detection and an enhanced focus on personalized primary prevention. �
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
�
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download �
* All of the above XYMOGEN policies remain strictly in force.
As a doctor who practices functional medicine, Dr. Jimenez utilizes the Neural Zoomer Plus. This is a blood test that analyzes neurological autoantibodies which offer very specific antibody-to-antigen recognition. The Neural Zoomer Plus tests the reactivity an individual has to 48 neurological antigens. These neurological antigens may be related to neurological disease and can help individuals assess the presence of a neurological condition.
There are 48 markers that are measured and they can be summed up and categorized into 7 larger groups. These groups include demyelination antigens, blood-brain barrier disruption, optical and autonomic nervous system disorders, peripheral neuropathy, neuromuscular disorders, brain autoimmunity, brain inflammation, and infections. (For a full list of the markers that the Neural Zoomer Plus measures, click here).�
The truth of the matter is that autoimmune disorders affect 5-10% of the general population and can target virtually and structure within the central or peripheral nervous system. Symptoms of an autoimmune disorder involving the CNS/PNS include but are not limited to:�
Having the tools to prevent a disease or disorder can be life-changing. With the ability to assess these markers, the rate of cognitive decline can steadily reduce. If you suffer from any of the above symptoms, the Neural Zoomer Plus may be right for you.
The intention of having our patients complete a Neural Zoomer Plus is to help us detect an individual’s IgA, IgG, and IgM sensitivity to antigens, down to the peptide level. Once we receive the results, not only do we have a resource that aids in the early detection of neurological diseases, but we also have a path. This path allows us to create a personalized prevention plan that will focus on the patient along with their lifestyle. – Kenna Vaughn, Senior Health Coach�
The Neural Zoomer Plus is a blood test that is designed to test 48 neurological antigens. When testing for these antigens, the results these markers find can help physicians determine if a patient is at risk for neurological conditions later on. To view last week’s article containing a full list of the signs and 48 markers, click here.�
When a patient comes to us with concerns, we listen very intently and make sure our patient’s concerns are addressed. More often than not, patients reveal they are having issues that relate to neurological declines, such as, muscle spasms or memory loss. With symptoms like these, the patient is referred to get a Neural Zoomer Plus.�
Once we receive the results back, it is compiled into a large report. From here, we assess it and go through all of the markers with an additional team of clinicians. An example of a few of the markers tested in a Neural Zoomer Plus is below. One can see that this patient has an elevated �Anti-Voltage gated potassium channel�. Anti- Voltage-gated potassium channels are responsible for multiple cellular processes such as cell growth and differentiation.�
After analyzing the data, we take these findings and create a treatment protocol that is specific to each patient and their lifestyle. Due to the fact that this patient shows an elevation in specific markers, there are certain adjustments we make in order to help prevent or reverse the cognitive and physical effects of neurological disorders.�
The first step we take is to change the diet of the patient. Many foods are not properly digested, leading to gut inflammation, which further leads to �leaky gut� which then enters the bloodstream and into the blood-brain barrier, causing neurological decline. In order to reverse these effects, we want to make sure the gut is getting the proper nutrients from foods that will not cause inflammation. For this patient, we recommend the Wahls Protocol.�
The second step we take is to usually get the patient involved in a more active lifestyle. By having this patient start to exercise with activities like yoga, it can improve the state of mind and their mind-body connection.�
The third step is usually nutraceuticals. These are supplements that will naturally help the body and brain with no addictive or harsh chemicals. With every patient, the nutraceuticals and amount needed vary depending on their specific body. For this case, we recommend:��
N-acetyl-L-cysteine:� (NAC) is a precursor to glutathione, the body�s most important cellular�antioxidant. NAC supplements have been shown to increase cellular glutathione levels.
Vitamins B12, B6, and folate: These are metabolic cofactors important for cellular metabolism and maintenance of all tissue cell types, but particularly important to nerve cells. Deficiencies in�B12 or folate can raise homocysteine levels, which have been associated with a higher risk for�vascular disease and dementia.
Alpha Lipoic Acid:� (ALA) is an essential cofactor in normal cellular metabolism and cellular�energy production.
Vitamin C and vitamin E: Can reverse symptoms caused by vitamin C and E�deficiencies.
As mentioned before, each patient is different and their lab work shows varying needs. However, with the Neural Zoomer Plus, we are able to get ahold of these symptoms, create a personalized treatment plan, and get them under control.�
As one can see, the data and knowledge we gain from these tests are truly eye-opening and give us an early advantage to help reverse or aid in prevention methods. We take the needs and concerns of every patient very seriously and work extremely hard to figure out the right method of treatment for them. Our goal is to help ensure that this lifestyle change is as smooth and easy on the patient as possible so they can get back to enjoying the activities they love and spending time with loved ones. The transition into a new lifestyle can be stressful, but with the information we gain from the tests, the knowledge we use from the doctor, and the willingness to change from the patient, we are set up to be the best team you can have to get your life back! – Kenna Vaughn, Senior Health Coach�
Microglial cells make up about 10 to 15 percent of all the glial cells in the human body, which can be found in the central nervous system (CNS) and play a fundamental role in the human brain. Microglial cells are responsible for maintaining and regulating changes in the physiological and pathological condition of the CNS by changing their morphology, phenotype and function. In an average physiological state, the microglial cells are continuously in charge of controlling their environment. �
However, when the homeostasis of the brain is interrupted, the microglia change into an amoeba-like shape and become a phagocyte where they can actively reveal a variety of antigens. If the homeostasis interruption in the CNS continues, the microglial cells will then trigger at a much stronger state, which is known as microglial priming. Microglia are the “Bruce Banner” of the CNS. However, once they go into protective “Hulk” mode, primed microglia become much more sensitive to stimulation and they have a much stronger possibility of reacting to stimulation, even reacting towards normal cells. �
�
Microglial priming can become a double-edged sword. As a matter of fact, primed microglia are created from different phenotypes of microglia and the phenotypes are context-dependent, which means they are associated to the sequence and duration of their exposure to different varieties of stimulation in a variety of pathologies. In the article below, we will demonstrate the effect of microglial priming on the central nervous system (CNS), especially in neurological diseases. �
Role of Microglial Cells in the CNS
Microglial cells are commonly found in the central nervous system (CNS), where they are considered to be one of the most flexible types of brain cells. Microglial cells are created from precursor cells found within mesoderm bone marrow, or more specifically found in the mesodermal yolk sac, and they are divided in different densities throughout several regions of the brain. As mentioned above, microglia will remain in a dormant state when the homeostasis of the brain remains stable. �
Microglia have a small cell body and morphological branches which extend towards all directions to help maintain and regulate the overall function of the CNS. Changes in their microenvironment can trigger microglia into an “activated� state. Research studies have demonstrated that microglia play a fundamental role in brain development and a variety of functions, including synaptic pruning and clearing out cell debris. Moreover, microglia create an immune surveillance system in the human brain and control fundamental processes associated with a variety of pathologies, including the clearance and uptake of A? and abnormal tau protein as well as the production of neurotrophic factors and neuroinflammatory factors. �
Microglial Priming Overview
Microglial priming activates when continuous interruptions in the brain’s microenvironment trigger a much stronger microglial response compared to an initial interruption which simply triggers microglial activation. Primed microglia in the CNS are also much more sensitive to possibly minor stimulation. This increased response involves microglial proliferation, morphology, physiology, and biochemical markers or phenotype. However, these changes will ultimately promote an increase in cytokines and inflammation mediator production which can have a tremendous impact on synaptic plasticity, neuronic survival, individual cognitive and behavioral function. Below is an overview of the effects of microglial priming in the CNS. �
Mechanisms of Microglial Priming in the CNS
The microenvironment of the central nervous system (CNS), by way of instance, is one of the main factors which can affect the microglial cells. Increased oxidative stress, lipid peroxidation and DNA damage associated with brain aging can all commonly trigger microglial priming. Another common factor for microglial priming includes traumatic brain injury. Research studies have shown that traumatic CNS injury activates microglia as well as the development of primed microglia. �
Many research studies have also shown that both focal and diffuse traumatic brain injury increase inflammation in the brain associated with microglia and astrocytes. CNS infections can also trigger microglial priming where viruses are the main cause of CNS infection. Both DNA and RNA viruses can trigger microglial priming including microglia and astrocytes. Recent research studies have shown that complement dysfunction can change the expression of complement receptors and trigger microglial priming after continuous activation following a variety of functions, including synapse maturation, immune product clearance, hematopoietic stem/progenitor cells (HSPC) mobilization, lipid metabolism, and tissue regeneration. �
Moreover, research studies have shown that there is increased priming of the microglia in a variety of neurological diseases. By way of instance, microglial cells with a morphological phenotype are found in large numbers in the human brain. In the last several years, research studies have suggested that neuroinflammation can continuously activate the microglia and trigger microglial priming. Furthermore, all of the previously mentioned situations are closely associated with neuroinflammation. Research studies have also demonstrated that neuroinflammation, as well as microbial debris and metabolic effects, are associated with central sensitization in neurological diseases, such as fibromyalgia, also referred to as the “brain on fire”. �
In the context of the previous situations mentioned above, microglia are primed though a series of pro-inflammatory stimulation, such as lipopolysaccharide (LPS), pathogenetic proteins (e.g., A?), ?synuclein, human immunodeficiency virus (HIV)-Tat, mutant huntingtin, mutant superoxide dismutase 1 and chromogranin A. There is also a variety of signaling pathways and it is common for different types of cells to express special pattern recognition receptors (PRRs) which can affect inflammatory signaling pathways. By way of instance, several signaling pathways, known as pathogen-associated molecular patterns (PAMPs), which can commonly increase in infected tissue, could also control microbial molecules. �
Additionally, peptides or mislocalized nucleic acids identified as misfolded proteins through a series of pathways, known as danger-associated molecular patterns (DAMPs), can also cause microglial priming. Toll-like receptors (TLRs) and carbohydrate-binding receptors commonly function in these pathways. There are also many different receptors found in microglia, including triggering receptors expressed on myeloid cells (TREM), Fc? receptors (Fc?Rs), CD200 receptor (CD200R), receptor for advanced glycation end products (RAGE), chemokine receptors (CX3CR1, CCR2, CXCR4, CCR5, and CXCR3), which can be recognized and mixed in with other signaling pathways, although some pathways are still not clear. �
Consequences of Microglial Priming in the CNS
Microglia show a low rate of mitosis in their normal state and a high rate of proliferation after microglial priming, showing that the microglia have the ability to affect cell turnover and pro-inflammation stimulation. With continued stimulation, microglia activate from their resting state, changing into amoeboid microglial cells in morphology. However, the changes in the shape of the microglia cannot differentiate the characteristics of microglial activation and the function of primed microglia depends on their phenotypes which are associated with receptors and molecules which they create and recognize. �
The different types of tissue macrophages, under microenvironmental impetus, are able to differentiate M1 and M2 phenotypes. First, M1 polarization, also known as classical activation, ultimately needs interferon-? (IFN-?) to be mixed with TLR4 signaling which then causes the production of inducible nitric oxide synthases (iNOS), reactive oxygen species (ROS), proinflammatory cytokines, and finally, ultimately reduces the release of neurotrophic factors, ultimately causing inflammation with increased markers of main histocompatibility complex II (MHC II), interleukin-1? (IL-1?) and CD68. �
Moreover, M2 polarization, also known as alternative activation, is ultimately believed to be associated with tissue-supportive in the situation of wound healing, reducing inflammation and improving tissue repair of collagen form. They trigger in response to IL-4 and IL-13 in vivo. M2 polarization is characterized by the increased expression of neurotrophic factors, proteases, enzymes arginase 1 (ARG1), IL-10 transforming growth factor-? (TGF-?), scavenger receptor CD206 and coagulation factors as well as improving phagocytic activity. As a matter of fact, there are currently no clear boundaries between the two polarizations and the M1 phenotype shares many similar characteristics with the M2 phenotype. �
Another phenotype of primed microglia, known as acquired deactivation, has been recently discovered. This new phenotype overlaps with M2 and has the ability to improve anti-inflammatory and functional recovery. Additionally, a research study conducted ultra-structural analyses and identified a brand-new phenotype, known as �dark microglia�, which is rarely seen in the microglial cell’s resting state. Systemic inflammation triggers microglia into an activated state to promote cell and tissue recovery and achieve homeostasis. Microglial priming is ultimately the second interruption in the CNS microenvironment. �
The primed microglia is a double-edged sword for brain health. Many research studies in vivo and in vitro have shown that neurological diseases are associated with microglial activation. The inflammatory phenotypes of the microglia create neurotoxic factors, mediators and ROS which can affect the CNS. Primed microglia play a fundamental and beneficial role in neuronal regeneration, repair, and neurogenesis. Primed microglia are also much more sensitive and respond much stronger to brain injury, inflammation, and aging as well as increase the activation of microglial cells by switching from an anti-inflammation, potentially protective phenotype to a pro-inflammation destructive phenotype, as shown in (Figure 1). �
�
In the early stages of microglial priming, the ability and function to phagocytize cell debris, misfolded proteins, and inflammatory medium are increased where more protective molecules, such as IL-4, IL-13, IL-1RA, and scavenging receptors, are created. The changes can affect wound healing and damage tissue repairment, neuron protection, and homeostasis recovery. Classically activated microglia (M1) make up a large proportion of all microglia and promote an increased creation of neurotoxic factors, such as IL-1?, TNF-?, NO and H2O2 (6), where more microglia are primed immediately afterward. �
This increased and extended neuroinflammation caused by primed microglia can ultimately be associated with the development and clustering of the protein tau and A?. Furthermore, it can lead to loss of neurons as well as the decrease of cognitive function and memory, such as in Alzheimer’s disease. Although the mechanisms are not clear enough, people have reached an agreement that primed microglia cause a chronic proinflammatory response and a self-perpetuating cycle of neurotoxicity. And this is believed to be the key factor in brain health issues resulting in neurological diseases. �
Microglia are known as the protectors of the brain and they play a fundamental role in maintaining as well as regulating the homeostasis of the CNS microenvironment. Constant stimulation causes the microglia to trigger at a much stronger state, which is known as microglial priming. Microglial cells are the “Bruce Banner” of the CNS. However, once they go into protective “Hulk” mode, primed microglia become much more sensitive to stimulation and they have a much stronger possibility of reacting to stimulation, even reacting towards normal cells. �- Dr. Alex Jimenez D.C., C.C.S.T. Insight
Microglial cells make up about 10 to 15 percent of all the glial cells in the human body, which can be found in the central nervous system (CNS) and play a fundamental role in the human brain. Microglial cells are responsible for maintaining and regulating changes in the physiological and pathological condition of the CNS. The scope of our information is limited to chiropractic, musculoskeletal and nervous health issues as well as functional medicine articles, topics, and discussions. To further discuss the subject matter above, please feel free to ask Dr. Alex Jimenez or contact us at 915-850-0900 . �
Curated by Dr. Alex Jimenez �
Additional Topic Discussion: Chronic Pain
Sudden pain is a natural response of the nervous system which helps to demonstrate possible injury. By way of instance, pain signals travel from an injured region through the nerves and spinal cord to the brain. Pain is generally less severe as the injury heals, however, chronic pain is different than the average type of pain. With chronic pain, the human body will continue sending pain signals to the brain, regardless if the injury has healed. Chronic pain can last for several weeks to even several years. Chronic pain can tremendously affect a patient’s mobility and it can reduce flexibility, strength, and endurance.
Neural Zoomer Plus for Neurological Disease
Dr. Alex Jimenez utilizes a series of tests to help evaluate neurological diseases. The Neural ZoomerTM Plus is an array of neurological autoantibodies which offers specific antibody-to-antigen recognition. The Vibrant Neural ZoomerTM Plus is designed to assess an individual�s reactivity to 48 neurological antigens with connections to a variety of neurologically related diseases. The Vibrant Neural ZoomerTM Plus aims to reduce neurological conditions by empowering patients and physicians with a vital resource for early risk detection and an enhanced focus on personalized primary prevention. �
Formulas for Methylation Support
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
�
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download�
* All of the above XYMOGEN policies remain strictly in force.
IFM's Find A Practitioner tool is the largest referral network in Functional Medicine, created to help patients locate Functional Medicine practitioners anywhere in the world. IFM Certified Practitioners are listed first in the search results, given their extensive education in Functional Medicine




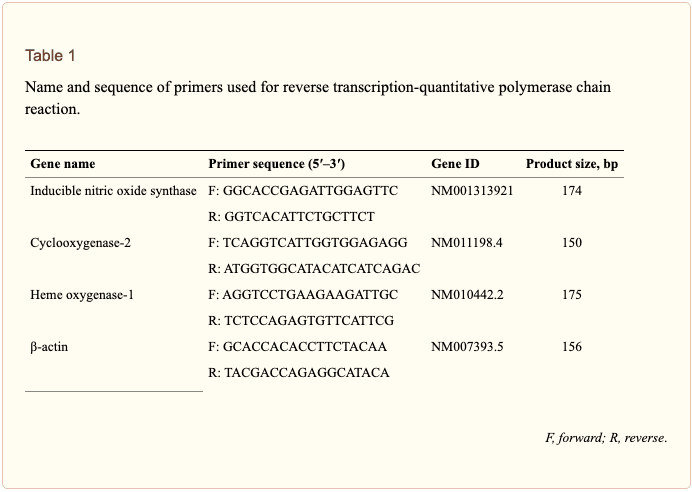
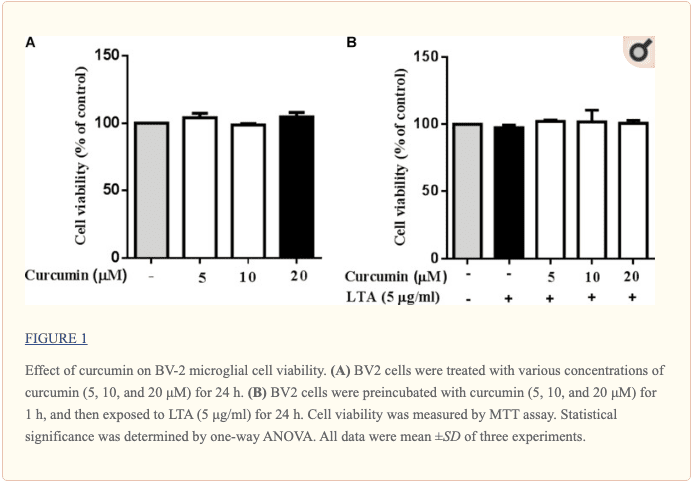
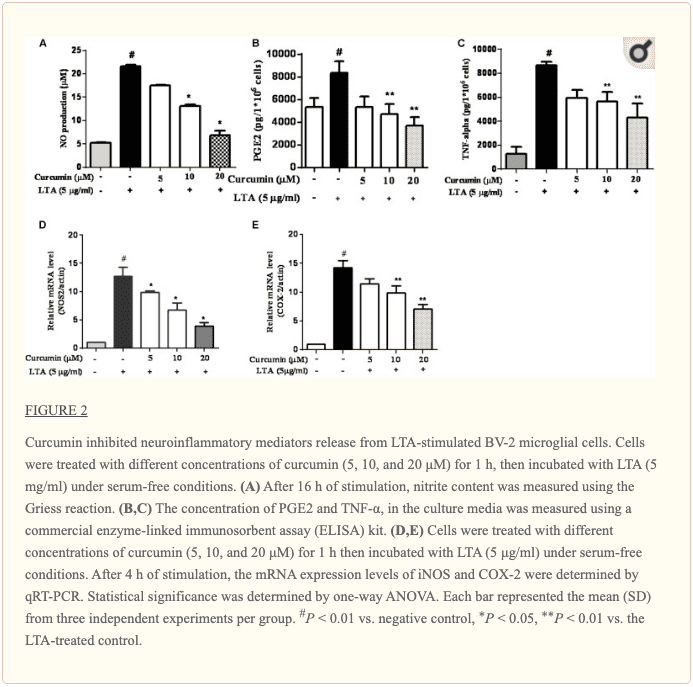
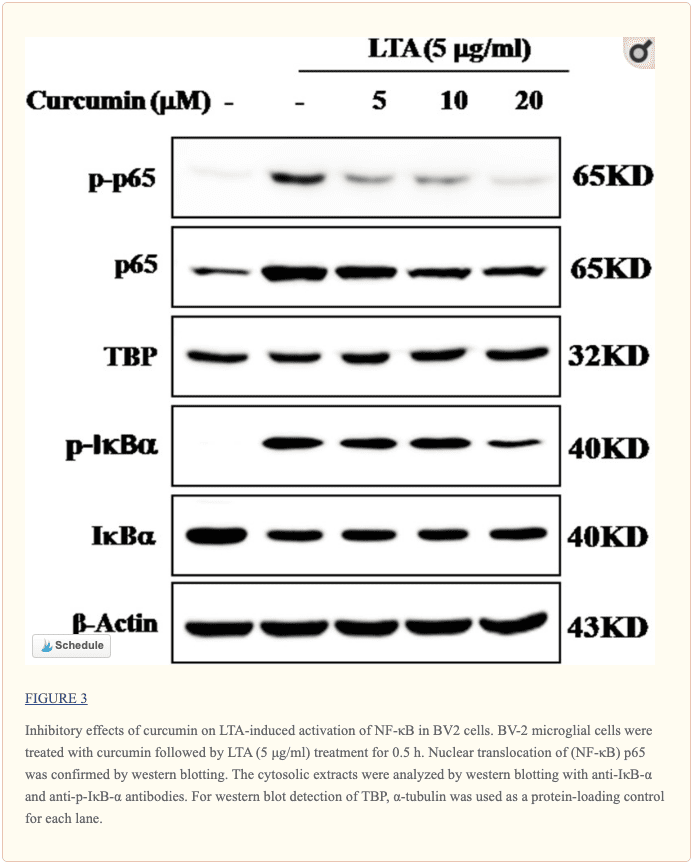
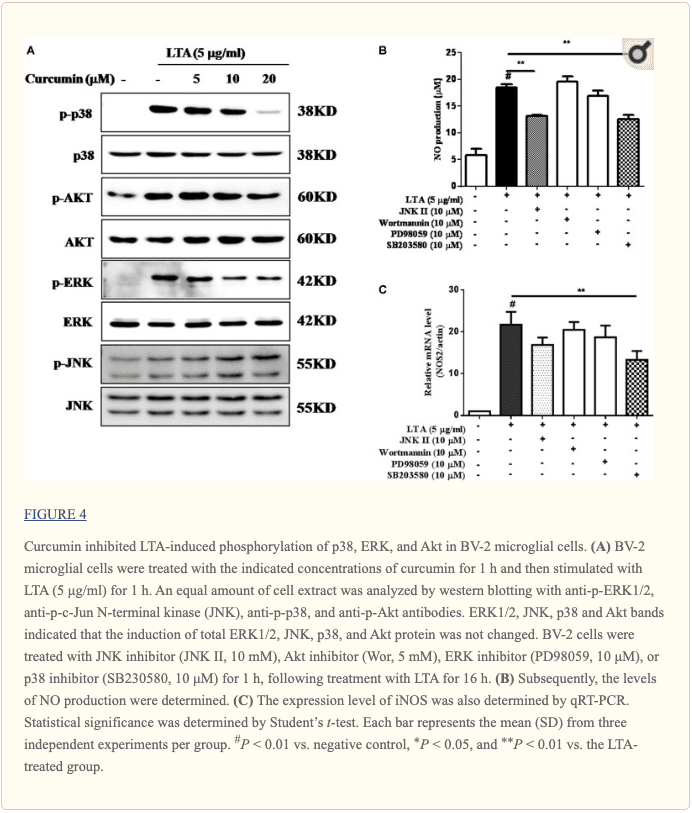
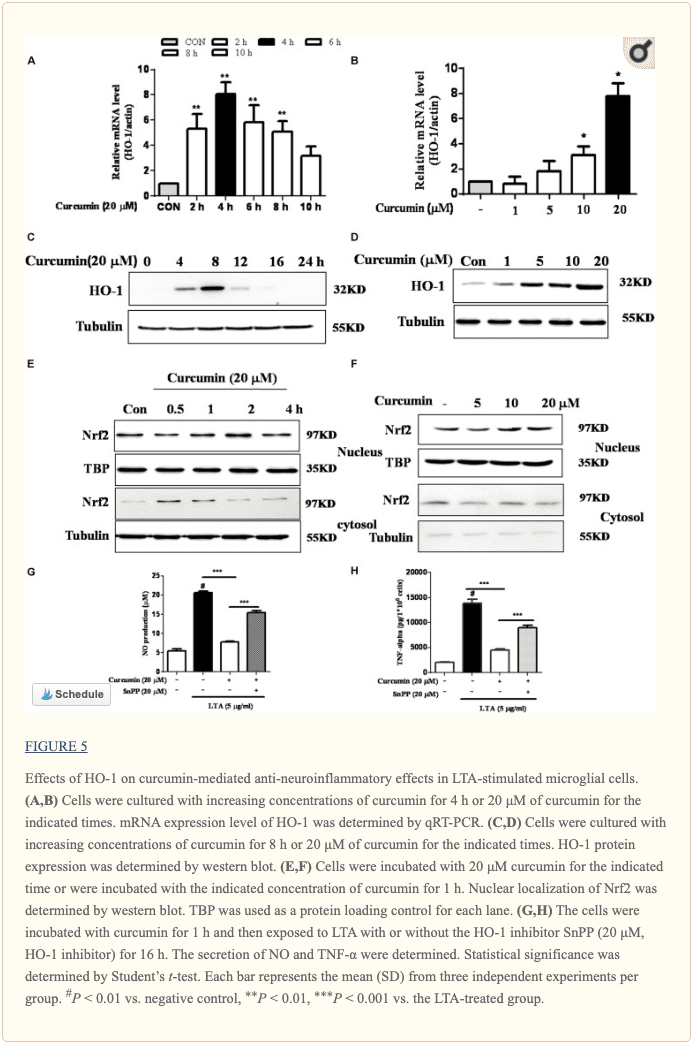
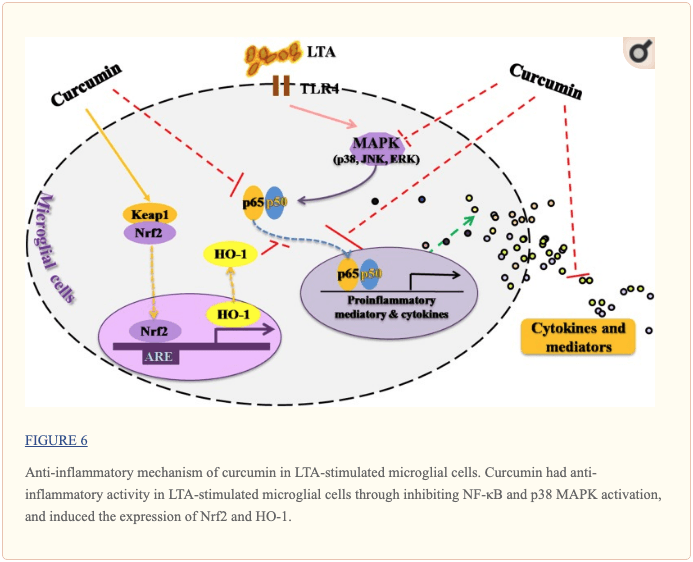 �
� 

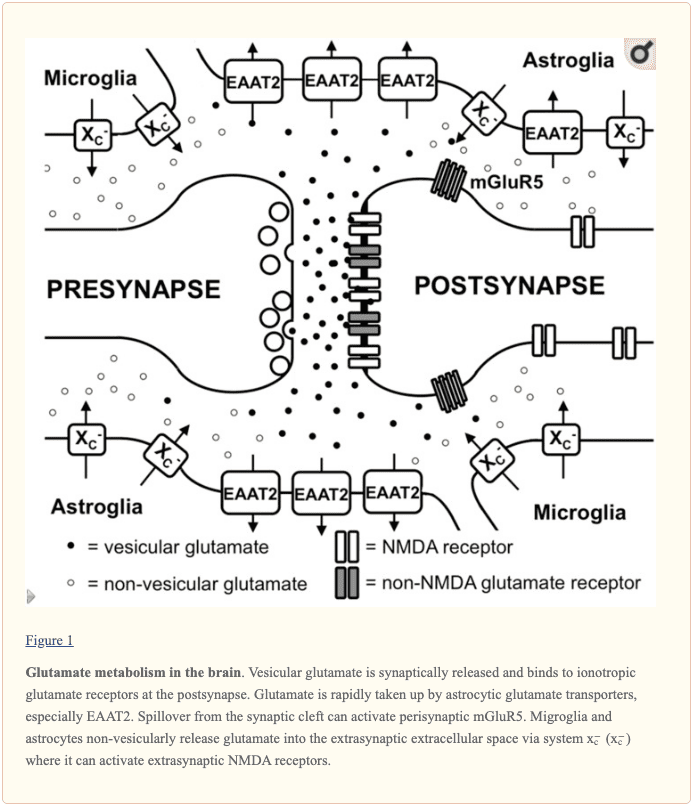


 �
�


 �
�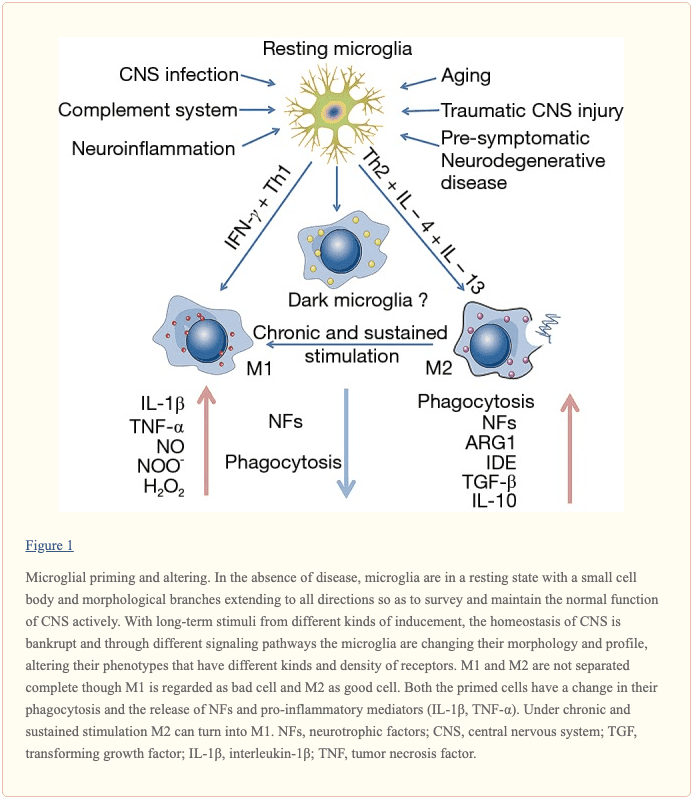 �
�






