Doctor de Medicina Funcional Explica los Signos Vitales
Hoy comenzaremos a analizar c�mo el espectro que la medicina actualmente considera “normal” puede no ser realmente �ptimo para su salud y bienestar general. Estos rangos de referencia pueden cambiar seg�n la edad, el sexo, la actividad f�sica y m�s. De hecho, si evalu�ramos el peso de una persona en los Estados Unidos, se considerar�a “normal” tener sobrepeso, simplemente porque el 70 por ciento de la poblaci�n tiene sobrepeso. Los rangos de referencia para las pruebas de laboratorio de hoy se basan en una poblaci�n enferma cuando debemos aspirar a un bienestar �ptimo.
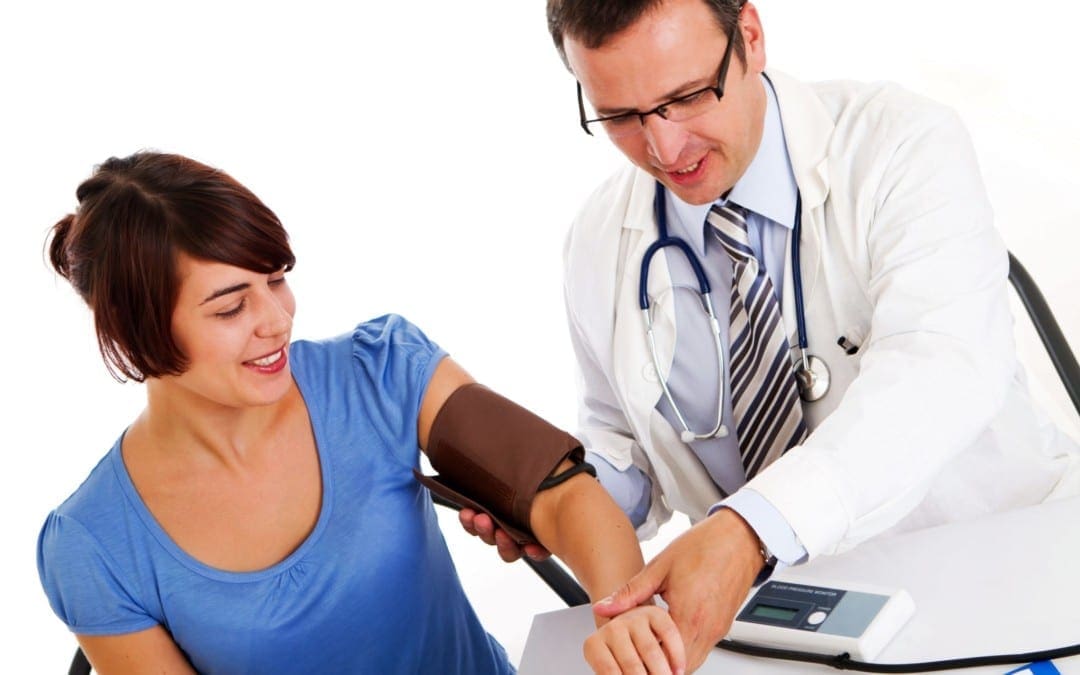
Luego, les demostrar� c�mo este conocimiento puede aplicarse a las medidas m�dicas m�s b�sicas: sus signos vitales. Todo el mundo sabe que cuando se visita a un m�dico por primera vez, toman sus signos vitales, incluyendo su peso, su presi�n arterial, su frecuencia card�aca y su temperatura. Sin embargo, �su m�dico le dice lo que significan sus resultados? �C�mo puede saber si est� sano?
�Cuales�son�los�Signos�Vitales?
Hola a todos, soy el Dr. Alex Jim�nez. Bienvenidos a la parte 2 de “C�mo Tomar el Control de su Salud”. Hoy, discutiremos los rangos de referencia de laboratorio asociados con sus signos vitales. Puede que no parezca un tema interesante, pero es realmente importante para su bienestar.
La mayor�a de los m�dicos generalmente definen los resultados de las pruebas de un paciente como “normales” o “anormales”. Pero, �qu� significa exactamente “normal” y “anormal”? Normal es cuando el 95 por ciento de la poblaci�n cae dentro de un rango com�n, mientras que anormal es cuando el porcentaje restante de la poblaci�n cae fuera de ese rango com�n. Ya sea que est� sano o enfermo, joven o viejo, aunque puede haber algunas variaciones para los ni�os, normal y anormal son simplemente n�meros estad�sticos que caen dentro de dos desviaciones est�ndar.
Sin embargo, estas desviaciones est�ndar no necesariamente significan que sea �ptimo o no. Es s�lo un n�mero estad�stico, despu�s de todo. De hecho, la enfermedad puede ocurrir en personas sanas. Estos rangos de referencia de laboratorio deben definirse de acuerdo con lo que es mejor para un ser humano.
A modo de ejemplo, los niveles de vitamina D se clasifican como normales si son m�s de 20, sin embargo, los niveles ideales son m�s de 50. �Entonces por qu� se considera que 20 es “normal”? Esto se debe a que aproximadamente el 80 por ciento de la poblaci�n es deficiente en vitamina D, por lo tanto, se encuentran dentro de lo que se considera el rango “normal”. Sin embargo, esto no significa que estos niveles sean los mejores para su salud y bienestar en general. Adem�s, los niveles “normales” de az�car en la sangre se han clasificado en menos de 100, aunque sabemos que los niveles “�ptimos” de az�car en la sangre se han clasificado entre 70 y 80. Los niveles de az�car en la sangre mayores de 80 pueden tener un mayor riesgo de enfermedad. Y desafortunadamente, nuestros rangos de referencia de laboratorio “normales” no son �ptimos porque nos hemos convertido en una poblaci�n enferma. En los Estados Unidos puede considerarse “normal” tener sobrepeso porque el 70 por ciento de nuestra poblaci�n tiene sobrepeso. Pero, aunque el sobrepeso u obesidad se considera normal en los Estados Unidos, no es algo a lo que quisi�ramos aspirar. Queremos aspirar a alcanzar la salud y el bienestar general.
Ahora continuemos discutiendo el significado de lo que es normal. Muchos pacientes a menudo visitan al m�dico solo para que se les diga que sus an�lisis de laboratorio han regresado normales, sin embargo, pueden sentirse enfermos. Qu� significa eso? �Significa que est� enfermo? �Significa que est� sano? Como mencion� anteriormente, o a su m�dico le falta algo o usted est� loco, pero estoy bastante seguro de que a su m�dico le falta algo. Esta es una de las principales diferencias entre la medicina convencional y la medicina funcional. A trav�s de la medicina funcional, muchos m�dicos se enfocan en el cuidado de la salud en lugar del cuidado de la enfermedad. Estamos buscando desviaciones m�s sutiles de las �ptimas.
Hasta que su funci�n se considere anormal seg�n el est�ndar actual, es posible que sus c�lulas ya est�n muriendo. Un m�dico de medicina funcional puede revisar sus pruebas de laboratorio de manera diferente a un m�dico convencional. Esto se debe principalmente a que los rangos de referencia en los que nos enfocamos apuntan hacia una salud �ptima, no a la enfermedad. Muchos m�dicos convencionales eval�an las pruebas de laboratorio de manera diferente a los m�dicos de medicina funcional, y luego siguen un “enfoque de espera”, o lo etiquetan como “no enfermo” despu�s de la cantidad m�nima de pruebas de laboratorio. De hecho, un paciente que me visit� ten�a niveles de az�car en la sangre de 120, donde un nivel de az�car en la sangre de 126 ya se considera diabetes tipo 2. Y le dije: “�Visito a su m�dico con respecto a esto?” Y �l dijo: “S�”. Le pregunt� sobre qu� le dec�a el doctor. Y finalmente dijo: “Bueno, �l dijo que esperara hasta que yo realmente tuviera diabetes y luego volviera por medicamentos”. Y eso es lo �ltimo que queremos hacer como profesionales de la salud.
Los rangos de referencia nos dan un n�mero promedio de valores que se han registrado entre la poblaci�n general. Pero no olvidemos que los rangos de referencia “normales” son relativos. Cambian seg�n la edad, el g�nero, la actividad f�sica y m�s. Si tuviera que evaluar el peso de una persona en los Estados Unidos hoy, ser�a normal tener sobrepeso, como lo mencion�, solo porque el 70 por ciento de las personas tienen sobrepeso. Y desafortunadamente, seguimos cambiando nuestros rangos de referencia en funci�n de nuestra poblaci�n enferma. Esto no es lo que deber�amos aspirar a hacer como m�dicos. Es por esto que la medicina funcional trata al individuo, no solo a los n�meros.
Adem�s, los rangos de referencia que alguna vez se consideraron normales tambi�n pueden cambiar con el tiempo. Un ejemplo de c�mo cambian los rangos de referencia fue demostrado por una compa��a de laboratorio global conocida como LabCorp, donde recientemente cambiaron sus rangos de referencia para los niveles de testosterona masculina. Anteriormente, LabCorp consideraba que los niveles normales de testosterona para un hombre adulto estaban entre 348 y 1.197. Este valor se bas� en una poblaci�n de machos adultos saludables. Sin embargo, en 2017, bajaron los niveles normales de testosterona para un hombre adulto entre 264 y 916. Adem�s, los hombres con sobrepeso, excluyendo a los hombres obesos, probablemente se incluyeron en el estudio, lo que finalmente cambi� los rangos de referencia para los niveles de testosterona masculina. Los estudios de investigaci�n han encontrado que el exceso de grasa abdominal puede causar niveles m�s bajos de testosterona. Sin embargo, al cambiar los rangos de referencia, esto demuestra que la medicina convencional considera que los individuos con sobrepeso son parte de la norma. Pero esto no es lo que queremos. Queremos luchar por el bienestar general.
Es por esto que necesita comenzar a tomar control de su propia salud. Como una de cada dos personas tiene alg�n tipo de enfermedad cr�nica, tenemos que evaluar c�mo interpretamos las pruebas de laboratorio y c�mo �normal� puede no significar necesariamente salud y bienestar, sino simplemente un promedio para una poblaci�n enferma en crecimiento en los Estados Unidos.
Tomando el Control de sus Signos Vitales
El objetivo principal de esta serie de videos es alentarlo a que se convierta en el l�der de su propio bienestar al comprender lo que significan sus ex�menes de laboratorio, a comprender c�mo se ve el aspecto �ptimo y al comprender qu� ex�menes de laboratorio est�n dise�ados para ayudarlo a lograr una salud general y bienestar en lugar de centrarse en la enfermedad. Tambi�n me gustar�a educarlo para que pueda tomar una decisi�n informada sobre a qui�n elige para ser su m�dico o compa�ero en su viaje hacia el bienestar. Ahora veamos las medidas m�dicas m�s comunes: sus signos vitales.
Sus signos vitales son tomados inicialmente por la enfermera cuando visita al m�dico. Estos signos vitales generalmente incluyen presi�n arterial, peso, frecuencia card�aca, temperatura e incluso saturaci�n de ox�geno. Sin embargo, �esta consciente de lo que significan estos n�meros? �Ha discutido su m�dico estos n�meros con usted? �Por qu� tomar�an sus signos vitales si solo los van a apuntar y nunca los discutir�n con usted? �Los n�meros realmente demuestran su salud y bienestar? Si tiene presi�n arterial alta o un problema de ritmo card�aco, lo m�s probable es que su profesional de la salud le avise, pero de lo contrario, es posible que no descubra cu�l es el valor de sus signos vitales.
Su frecuencia card�aca es probablemente uno de los signos vitales m�s importantes que se toman durante una visita al m�dico. El pulso es una medida de qu� tan r�pido est� latiendo su coraz�n. El coraz�n humano late m�s de 115,000 veces por d�a. Entonces, si su frecuencia card�aca est� por encima de 100, entonces definimos eso como tener una frecuencia card�aca alta. Pero, si tiene una frecuencia card�aca superior a 80, eso puede aumentar su riesgo de desarrollar una enfermedad card�aca. �Qu� causa este aumento? Aunque muchos factores pueden conducir a una enfermedad cardiovascular, el estr�s es una de las causas m�s comunes porque aumenta la adrenalina y causa un aumento del ritmo card�aco y la presi�n arterial. Tomar demasiado caf�, medicamentos estimulantes como Adderall o un problema de salud de la tiroides, y el coraz�n o los pulmones hiperactivos, tambi�n pueden aumentar el riesgo de enfermedades del coraz�n.
La respuesta de lucha o escape, una reacci�n fisiol�gica que se activa en momentos de estr�s, tambi�n puede causar un aumento en la frecuencia card�aca. Cuando el ritmo card�aco de un individuo con frecuencia es superior a 80, puede ser el momento de comenzar a incorporar algunas t�cnicas de manejo del estr�s en su vida, por ejemplo, la meditaci�n consciente y otras formas de meditaci�n. El estr�s no es la �nica causa de un aumento de la frecuencia card�aca. La ansiedad, las deficiencias de magnesio, la falta de condicion y la deshidrataci�n, tambi�n pueden causar un aumento de la frecuencia card�aca. Idealmente, queremos lograr una frecuencia card�aca m�s baja, �ptimamente por debajo de 70.
En el otro lado del espectro, una disminuci�n de la frecuencia card�aca por debajo de 60 tambi�n podr�a demostrar la presencia de disfunci�n de la tiroides o una funci�n tiroidea baja. Los atletas y los corredores de distancia en realidad tienen una frecuencia card�aca baja porque est�n muy bien acondicionados. Su frecuencia card�aca puede ser tan baja, como 50 o incluso 45. Pero, si tiene una frecuencia card�aca baja y no es un atleta o un corredor de distancia, puede que sea el momento de hablar con su m�dico.
Aunque la frecuencia card�aca es uno de los signos vitales m�s importantes que se toman durante una visita al m�dico, existe otro signo vital que puede ser igual de importante: la variabilidad de la frecuencia card�aca. Esto refleja la salud de su sistema nervioso autom�tico o aut�nomo, que se encarga de controlar todos los elementos inconscientes de su sistema nervioso, como la digesti�n y la respiraci�n. La variabilidad de la frecuencia card�aca se ha asociado frecuentemente con la longevidad e incluso la muerte. Cuanto menos variable sea la frecuencia card�aca, mayor ser� la tasa de mortalidad. Muchos m�dicos no miden la variabilidad del ritmo card�aco de un paciente, pero afortunadamente, usted puede controlarlo por si mismo. Las terapias de fr�o y calor, los saunas, el ejercicio, el yoga y la meditaci�n pueden ayudar a mejorar la variabilidad del ritmo card�aco del paciente.
Ahora pasemos a la siguiente, la m�s importante de los signos vitales tomados durante una visita al m�dico, la presi�n arterial. Si tuviera que alinear todos los vasos sangu�neos de su cuerpo, se extender�an aproximadamente 59,000 millas. Eso es casi siete veces alrededor de la tierra. Estos mismos vasos sangu�neos transportan m�s de 7,500 litros de sangre por todo el cuerpo de manera regular. Con cada latido del coraz�n, la sangre se empuja contra las paredes de las arterias, lo que provoca un aumento de la presi�n.
Las medidas m�dicas para la presi�n arterial tienen dos n�meros. El n�mero superior se conoce como sist�lica, o la presi�n cuando el coraz�n se est� contrayendo, y el n�mero inferior se conoce como diast�lica, o la presi�n cuando el coraz�n est� relajado o en reposo. Los rangos de referencia normales para la presi�n arterial contin�an cambiando porque seguimos descubriendo que los rangos de referencia que sol�amos considerar normales, que fueron primero 140/90, luego 130/80, todav�a estaban asociados con un mayor riesgo de ataque cerebral y apoplej�a . Hoy en d�a, muchos m�dicos pueden mencionar un problema con su presi�n arterial solo si tiene m�s de 130/80.
La raz�n por la que la presi�n arterial es tan importante es porque cuando est� elevada, puede ejercer una presi�n adicional sobre el coraz�n y las arterias, lo que podr�a provocar una enfermedad card�aca, un accidente cerebrovascular, una insuficiencia card�aca o incluso una insuficiencia renal. Y no solo el coraz�n y las arterias se ven afectados por la presi�n arterial: el cerebro, los ri�ones e incluso los ojos pueden verse tremendamente afectados, lo que provoca accidentes cerebrovasculares, demencia, insuficiencia renal y ceguera, entre otros problemas de salud. Mantener su presi�n arterial a un nivel �ptimo es fundamental para su salud y bienestar general. De hecho, actualmente se cree que la presi�n arterial normal es inferior a 120/80, sin embargo, puede llegar a ser incluso m�s baja.
Mientras que la presi�n arterial alta es mala, la presi�n arterial baja tambi�n puede ser mala. Un buen m�dico de medicina funcional discutir� con usted los riesgos tanto de la presi�n arterial alta como de la presi�n arterial baja. La presi�n arterial por debajo de 100/60 puede causar problemas, pero no necesariamente. Los vasos sangu�neos en el cuerpo humano funcionan como los pistones en un autom�vil. Si no se acumula suficiente presi�n, puede ser realmente dif�cil que la sangre fluya contra la gravedad. Y debido a que el cerebro humano est� en una posici�n m�s alta que el coraz�n, dependemos de nuestra presi�n arterial para suministrar a nuestro cerebro la cantidad necesaria de ox�geno y nutrientes necesarios para funcionar en consecuencia.
Si tiene presi�n arterial baja, puede experimentar otros s�ntomas, como fatiga. Otros s�ntomas asociados con la presi�n arterial baja incluyen mareos al estar de pie, debilidad e incluso niebla cerebral. Adem�s, tanto la presi�n arterial alta cr�nica como la presi�n arterial baja cr�nica pueden contribuir a un mayor riesgo de demencia.
Ahora hemos discutido la importancia de la frecuencia card�aca y la presi�n arterial. Pero, �Qu� tal si hablamos de otro signo vital importante: la temperatura de su cuerpo? Una fiebre o una temperatura corporal elevada a menudo pueden ser un signo de infecci�n. La temperatura tambi�n puede proporcionar una visi�n de la funci�n de nuestro metabolismo. Cuanto m�s bajo es el metabolismo de un individuo, menos calor produce, que puede manifestarse como una temperatura corporal ligeramente m�s baja de lo normal. La tiroides juega un papel importante en el metabolismo y en la regulaci�n de la temperatura. Por lo tanto, si con frecuencia se siente fr�o, es posible que desee hablar con su m�dico sobre c�mo solicitar el panel de tiroides correcto. Pero, �qu� pruebas debe pedirle a su m�dico para determinar esto? No se preocupe, analizaremos qu� pruebas de tiroides debe realizar en el video sobre las hormonas. La temperatura corporal �ptima debe ser de aproximadamente 98.6 grados Fahrenheit. Sin embargo, si es inferior a 97.7 grados Fahrenheit, puede indicar que tiene un problema de tiroides.
Las medidas m�dicas finales que vamos a discutir son su estatura y su peso. Los m�dicos utilizan su altura y peso para calcular su �ndice de masa corporal o IMC. En nuestra cl�nica, a modo de ejemplo, utilizamos el InBody 770, un analizador de composici�n corporal y agua corporal, para ayudar a determinar f�cilmente su �ndice de masa corporal y m�s. Sin embargo, el �ndice de masa corporal no siempre tiene en cuenta la composici�n corporal o el porcentaje de grasa contra m�sculo en el cuerpo humano. A modo de ejemplo, un jugador de f�tbol profesional de 6’6 “y 265 libras tiene un �ndice de masa corporal de m�s de 30, lo que los coloca en la categor�a de obesos.
Pero si viera el cuerpo de este individuo, nunca se clasificar�an como obesos. Esto demuestra que el IMC no es una medida precisa, especialmente para los atletas. Adem�s, una mujer de 65 a�os puede tener m�s grasa que m�sculo en su cuerpo, mientras que sus medidas de IMC pueden parecer “�ptimas”. En cambio, muchos m�dicos de medicina funcional usan medidas m�dicas de cintura a cadera. Esta es una medida simple que puede hacer en casa para determinar la distribuci�n de la grasa corporal, que tambi�n puede ayudar a demostrar el riesgo de disfunci�n metab�lica. La obesidad, la diabetes tipo 2 y las enfermedades del coraz�n son causadas por el exceso de grasa abdominal o grasa acumulada alrededor de los �rganos. El exceso de grasa alrededor de la secci�n media puede, en �ltima instancia, aumentar el riesgo de enfermedades card�acas y problemas metab�licos, como diabetes, demencia, c�ncer y muchos otros problemas de salud.
Pero primero, hablemos de c�mo puede calcular su relaci�n cintura-cadera. Para medir su cintura, simplemente tome las medidas del �rea m�s ancha alrededor de su cintura, que generalmente es la parte m�s grande alrededor de su ombligo. Para medir su cadera, luego tome las medidas del �rea m�s ancha alrededor de su cadera, que es generalmente donde sus huesos de cadera est�n de los lados. Entonces, toma estas medidas y luego divide las medidas de su cintura por las medidas de su cadera. Y este es el n�mero m�s fundamental que tiene que mirar.
En los hombres, una relaci�n cintura-cadera inferior a 0,9 se considera �ptima. Si la proporci�n es mayor que uno, lo que significa que su vientre es m�s grande que su cadera, puede poner a los hombres en mayor riesgo de desarrollar s�ndrome metab�lico, enfermedad card�aca, diabetes, accidente cerebrovascular, c�ncer y demencia. En las mujeres, una proporci�n de cintura-cadera de menos de 0,8 se considera �ptima. Si la proporci�n es mayor que 0,85, puede poner a las mujeres en mayor riesgo de desarrollar s�ndrome metab�lico, as� como a los otros problemas de salud mencionados anteriormente.
Las medidas m�dicas, incluyendo la frecuencia card�aca, la temperatura, la frecuencia respiratoria y la presi�n arterial, son varios signos vitales que ayudan a indicar a los m�dicos el estado de las funciones corporales fundamentales de un paciente. Los rangos de referencia de hoy en d�a se utilizan para determinar los espectros “normales” de salud y bienestar, sin embargo, los estudios de investigaci�n han demostrado que estos rangos de referencia pueden no ser espectros �ptimos. Comprender las medidas m�dicas m�s b�sicas, o los signos vitales, es importante para el bienestar del paciente, ya que puede ayudar a las personas a reconocer si se sienten saludables o enfermos, independientemente de lo que es normal. Dr. Alex Jimenez D.C., C.C.S.T.
Entendiendo sus Signos Vitales
Y estos fueron sus signos vitales, sus medidas m�dicas m�s b�sicas. Estos n�meros son muy importantes ya que son fundamentales para su salud y bienestar en general. Comprender c�mo funciona su cuerpo como un todo es importante para optimizar su bienestar. Por lo tanto, la pr�xima vez que visite a su m�dico, pregunte sobre sus signos vitales y discuta estos rangos de referencia con ellos. Realmente creo que una combinaci�n de su propia investigaci�n y tener una buena relaci�n con un profesional de la salud calificado puede llevarlo por el camino correcto hacia la salud y el bienestar en general.
Encontrar un m�dico que trabaje con usted es esencial para lograr los resultados que merece. Si su m�dico no est� dispuesto a conversar con usted sobre su bienestar o las pruebas de laboratorio que se necesitan para realizar un seguimiento de sus resultados, es posible que desee considerar la posibilidad de buscar otro m�dico.
Si aprendi� acerca de las medidas m�dicas m�s simples, definitivamente disfrutar� el pr�ximo articulo, donde analizaremos los an�lisis de sangre utilizados para determinar las deficiencias nutricionales. M�s del 90 por ciento de las personas en los Estados Unidos son deficientes en nutrientes a nivel de la RDA. Esa es la cantidad m�nima necesaria para prevenir enfermedades causadas por deficiencias nutricionales.
Vamos a discutir c�mo un m�dico que se especializa en medicina funcional eval�a los resultados y qu� otras pruebas que su m�dico desconoce pueden decirnos mucho sobre su estado nutricional. Gracias de nuevo por acompa�arme hasta ahora y los ver� m�s tarde. El alcance de nuestra informaci�n se limita a problemas quiropr�cticos y de salud de la columna, as� como a temas y discusiones de medicina funcional. Para seguir discutiendo el tema, no dude en preguntarle al Dr. Alex Jimenez o comun�quese con nosotros al�915-850-0900�.
Curado por el Dr. Alex Jim�nez
Discusi�n del Tema Adicional: Dolor de Espalda Agudo
El dolor de espalda es una de las causas m�s frecuentes de discapacidad y d�as perdidos en el trabajo en todo el mundo. El dolor de espalda se atribuye a la segunda raz�n m�s com�n para las visitas al consultorio del m�dico, superada �nicamente por infecciones respiratorias superiores. Aproximadamente el 80 por ciento de la poblaci�n experimentar� dolor de espalda al menos una vez a lo largo de su vida. La columna vertebral es una estructura compleja formada por huesos, articulaciones, ligamentos y m�sculos, entre otros tejidos blandos. Las lesiones y / o afecciones agravadas, como las hernias de disco, pueden provocar s�ntomas de dolor de espalda. Las lesiones deportivas o las lesiones por accidentes automovil�sticos suelen ser la causa m�s frecuente de dolor de espalda; sin embargo, a veces los movimientos m�s simples pueden tener resultados dolorosos. Afortunadamente, las opciones de tratamiento alternativo, como la atenci�n quiropr�ctica, pueden ayudar a aliviar el dolor de espalda mediante el uso de ajustes de la columna vertebral y manipulaciones manuales, lo que finalmente mejora el alivio del dolor. �
De XYMOGEN�Las f�rmulas profesionales exclusivas est�n disponibles a trav�s de profesionales de atenci�n m�dica con licencia seleccionados. La venta por internet y el descuento de f�rmulas XYMOGEN est�n estrictamente prohibidos.
Con orgullo�El Dr. Alexander Jimenez�hace que las f�rmulas de XYMOGEN est�n disponibles solo para los pacientes bajo nuestro cuidado.
Llame a nuestro consultorio para que podamos asignar una consulta m�dica para acceso inmediato.
Si eres paciente de�Injury Medical & Chiropractic�Clinic, puedes preguntar por XYMOGEN llamando�915-850-0900.
Para su conveniencia y revisi�n de la�XYMOGEN�productos por favor revise el siguiente enlace. *XYMOGEN-Descargar-Catalogo
* Todas las pol�ticas de XYMOGEN anteriores se mantienen estrictamente.
El Doctor de Medicina Funcional Explica el Cuidado de Enfermos y el Cuidado de Salud
Hoy analizaremos los fundamentos de la medicina funcional y c�mo puede construir una relaci�n saludable entre m�dico y paciente.
Si visita al m�dico porque ha experimentado migra�as, eczema, s�ndrome del intestino irritable y depresi�n, lo m�s probable es que lo refieran a cuatro especialistas diferentes e incluso se le receten un m�nimo de cuatro medicamentos diferentes. Un enfoque de la medicina funcional comprende que puede haber problemas comunes de salud que pueden estar causando los s�ntomas de un paciente. Una vez que llegue al origen del problema, se puede solucionar los problemas de salud que crean los s�ntomas.
La medicina funcional pregunta: “�Por qu� tiene esos s�ntomas y c�mo podemos tratar la fuente del problema y mejorar su salud y bienestar general?”, en lugar de “�Qu� enfermedad tiene y qu� medicamento usa para tratarla?”
�Que es el Cuidado de Enfermos y el Cuidado de Salud?
Hola a todos, mi nombre es Dr. Alex Jim�nez. Soy quiropr�ctico y practicante de medicina functional en El Paso, Texas. Estoy muy contento/a de poder presentarles la primera parte de “C�mo Tomar el Control de su Salud”, donde analizaremos las diferencias entre �el Cuidado de Enfermos y el Cuidado de la Salud”. Por lo tanto, con frecuencia escucho a las personas hablar sobre lo dif�cil que es encontrar al m�dico adecuado, a alguien que est� dispuesto a trabajar con ellos, que los escuche y que est� abierto a satisfacer sus demandas mientras les ense�a todo lo que necesitan saber sobre sus problemas. Un m�dico que acepta avances innovadores en la ciencia y los enfoques de sistemas para determinar la fuente del problema. Esto se conoce como medicina funcional, y discutiremos m�s de eso en un minuto.
Mi objetivo en este momento es mostrarle c�mo puede encontrar al m�dico adecuado y establecer la mejor relaci�n con ellos para obtener la atenci�n que merece. Es importante hacerle varias preguntas a su m�dico para determinar si son adecuados para usted, si est�n dispuestos a trabajar con usted, a escucharlo y si est�n abiertos a sus solicitudes, as� como a aprender sobre los problemas que se les presentan. En lugar de no est�r familiarizado con ellos o que no sigan nuevos enfoques de tratamiento si se les presenta. A modo de ejemplo, si el m�dico que elige dice que la nutrici�n no tiene nada que ver con una enfermedad, entonces probablemente deber�a buscar otro m�dico. Ahora hablemos sobre las preguntas que debe hacerle a su m�dico sobre su salud y bienestar general para estar informado sobre lo que est� sucediendo en su propio cuerpo.
He hablado con pacientes que han visitado a numerosos m�dicos con la esperanza de encontrar una respuesta sobre por qu� no se sienten bien o tienen alg�n tipo de enfermedad. Muchos son referidos frecuentemente a uno o m�s especialistas y, en algunos casos, reciben Prozac, o se les dice que todos sus s�ntomas est�n en su cabeza o a veces incluso se les dice que no hay nada de malo despu�s de que todos los resultados de su laboratorio son normales. Y es frecuente que estos mismos pacientes sean enviados a varios m�dicos por cada parte de su cuerpo en lugar de ser enviados a un m�dico que pueda diagnosticar y tratar su cuerpo por completo. Si acude al m�dico y tiene migra�as, eczema, s�ndrome del intestino irritable y depresi�n, en la mayor�a de los casos, se le enviar� a cuatro especialistas diferentes y se le administrar�n, al menos, cuatro medicamentos diferentes, en lugar de visitar a un especialista que pueda comprender la fuente subyacente del problema y tratar la ra�z de los s�ntomas.
�Alguna vez ha visitado a un m�dico que, despu�s de explicarles sus s�ntomas, dice: “Oh, hice un panel de sangre completo y todo regreso normal”. Esto podr�a significar una de dos cosas, ya sea que est� loco o que les esta faltando algo. Y estoy seguro de que a menudo falta algo porque no buscan las respuestas en los lugares correctos. Es como una persona que deja caer sus llaves en la calle y cuando su amigo lo ve buscando debajo de una farola, dice: “Oye, �qu� est�s haciendo?”, Dice: “Estoy buscando mis llaves”. Entonces dice, �Pues d�nde las perdiste “. Y el dice:” Bueno, las perd� por la calle “.”� Entonces por qu� est�s buscando aqu�? “” Bueno, la luz est� mejor aqu� “. Y desafortunadamente, eso pasa mucho en la medicina. Los m�dicos buscar�n respuestas donde los problemas son f�ciles de encontrar. Y ese es el prop�sito de este video. Para ayudarlo a comprender c�mo puede encontrar la fuente de los problemas de salud subyacentes que lo hacen sentir mal.
Esto me recuerda a un caso de una mujer que vi en mi consultorio que ten�a artritis psori�sica, una afecci�n autoinmune que le caus� erupciones en todo el cuerpo junto con dolor e inflamaci�n en las articulaciones. Como resultado, ella consum�a grandes cantidades de drogas y / o medicamentos que sumaban alrededor de $60,000 d�lares al a�o o m�s. Tambi�n ten�a otros problemas de salud, ten�a reflujo �cido, s�ndrome del intestino irritable, pre-diabetes, migra�as, insomnio y depresi�n. Esta mujer visit� a muchos especialistas diferentes y estaba tomando recetas para tratar cada uno de sus s�ntomas. Cuando lleg� el momento de verla, todo lo que hice fue llegar a la fuente del problema.
Entonces, durante el diagn�stico, me dije a m� mismo: “Bueno, esta paciente tiene inflamaci�n, pero �cu�l es la causa de su inflamaci�n?” Y en lugar de recomendarla a muchos otros especialistas para que pudiera recibir medicamentos para sus migra�as, reflujo �cido, depresi�n, etc., me dije a m� mismo: “Oh, todos estos s�ntomas son inflamatorios, entonces, �cu�l es la ra�z de la inflamaci�n?” Bueno, resulta que la paciente tuvo problemas con su intestino todo el tiempo. Luego la ayud� a eliminar los “bichos malos” en su intestine, recomend�ndole una dieta anti-inflamatoria, inclu� algunos suplementos, aceite de pescado, vitamina D, probi�ticos. Sinceramente, remedios naturales realmente b�sicos. Cuando regres� seis semanas despu�s, todos sus s�ntomas hab�an desaparecido. Se hab�a retirado todos sus medicamentos recetados y hab�a perdido 20 libras. No le dije que dejara de tomar sus medicamentos, simplemente lo hizo por su propia cuenta. Fue absolutamente extraordinario y eso es lo que sucede cuando se trata la fuente de los problemas de salud subyacentes de un paciente. Y no tiene que hacer mucho para llegar a la ra�z del problema.
Entiendo que es posible que no siempre se pueda trabajar con un m�dico que est� capacitado en medicina funcional, de lo que hablaremos en un minuto. Sin embargo, es posible encontrar un m�dico que est� dispuesto a trabajar con usted, qui�n va a escuchar, qui�n tendr� la mente abierta y qui�n ser� su compa�ero durante su viaje hacia la salud y el bienestar en general. A lo largo de este video, analizar� las pruebas de laboratorio convencionales y las pruebas de laboratorio de medicina funcional innovadora, demostrando todo con el prop�sito de crear bienestar en lugar de verlo todo desde la perspectiva de la enfermedad para encontrar la fuente de los problemas de salud de un paciente y corregir los desequilibrios a trav�s de un enfoque de medicina funcional.
Tomando el Control de el Cuidado de Enfermos y el Cuidado de Salud
La medicina funcional a menudo se conoce como el futuro de la medicina, pero muchos m�dicos la ofrecen actualmente si visita el lugar correcto. El prop�sito de la medicina funcional es diagnosticar y tratar la causa de una variedad de enfermedades mediante la evaluaci�n del cuerpo por completo, en lugar de analizar cada colecci�n de �rganos de forma independiente a trav�s de especialistas independientes. La medicina funcional trata todo el sistema, no solo los s�ntomas. Los m�dicos que practican la medicina funcional con frecuencia se preguntan: “�Por qu� tiene el paciente estos s�ntomas y c�mo puedo corregir las causas y mejorar su salud y bienestar general?”, En lugar de “�Qu� enfermedad tiene el paciente y qu� medicamento utilizan para tratarlo?�
He sido un quiropr�ctico practicante por m�s de 25 a�os. Y he presenciado innumerables milagros todos los d�as. Mis pacientes no solo encuentran alivio de sus s�ntomas, realmente logran alcanzar el bienestar general. Desafortunadamente, nuestro sistema de salud actual est� roto. Muchos m�dicos se ven tremendamente afectados debido a las limitaciones de tiempo establecidas por las compa��as de seguros para pagar las facturas, lo que hace que se apresuren a acudir a sus citas. A la vez, este ciclo continuo con frecuencia puede terminar dejando a los pacientes con preguntas sin respuesta y frustraci�n. Nuestro sistema de atenci�n m�dica actual hace que las personas se sientan impotentes, a menudo manteni�ndolas atrapadas con sus enfermedades.
Como parte de este sistema de salud, es importante que entendamos que algunas enfermedades no son tan f�ciles de tratar como un resfriado. Varias enfermedades tampoco se desarrollan de forma aleatoria. La mayor�a de las enfermedades que existen en la actualidad est�n relacionadas con su entorno y como estos factores externos e internos interact�an con sus genes y estilo de vida para influir en su salud y bienestar.
Lo que hoy conocemos como el enfoque de la �medicina convencional� se conoce generalmente como el juego de �nombrarlo, culparlo y domesticarlo�. Primero, el m�dico diagnosticar� al paciente y le proporcionar� una etiqueta, por ejemplo, dicen que un paciente tiene depresi�n. Ahora tienen el nombre de la enfermedad. Y finalmente, el m�dico tratar� la enfermedad con medicamentos recetados. Al final, el paciente se queda tomando un antidepresivo. Sin embargo, la depresi�n puede ser causada por una amplia gama de factores y la soluci�n no es necesariamente un antidepresivo. Este tipo de pr�ctica no solo est� desactualizada, sino que en realidad se considera bastante inseguro a largo plazo. Y a menudo no proporciona a los pacientes los resultados que necesitan. Las personas son diagnosticadas err�neamente y con frecuencia quedan enfermas sin recibir realmente la atenci�n adecuada que merecen.
La medicina convencional tambi�n puede ser �til en las etapas finales de algunas enfermedades, as� como para enfermedades agudas. Si tiene una emergencia o se siente muy enfermo, si se rompe un hueso, o si tiene una infecci�n, la medicina convencional, los cuidados intensivos, los medicamentos pueden ser la soluci�n adecuada y deber�amos estar sumamente agradecidos por ellos. Pero este no es el enfoque que necesitamos para prevenir y curar enfermedades cr�nicas. Si realmente entendi�ramos c�mo cuidar nuestros cuerpos como deber�amos, la mayor�a de nosotros no nos sentir�amos tan enfermos. Y muchas personas seguiran sinti�ndose enfermas, pero no tiene que hacerlo, no es normal. Muchos m�dicos ahora entienden que se requiere un cambio para convertir todo nuestro sistema de atenci�n de enfermos en uno que realmente respalde la atenci�n m�dica.
Me gustar�a capacitarlo para ayudar a transformar el futuro del campo de la medicina al tomar su bienestar en sus propias manos. A lo largo de los pr�ximos videos, compartir� con usted c�mo puede encontrar un m�dico que tenga en cuenta sus valores y creencias personales para que pueda lograr los resultados que est� buscando mientras se encuentra en un entorno seguro y c�modo. Puede aprender a ser su propio defensor de la salud y convertirse en un verdadero socio con su m�dico. Y hay muchos otros m�dicos y practicantes de medicina funcional como yo que est�n esperando para ayudar.
Adem�s, compartir� con usted c�mo puede encontrar al mejor m�dico y tambi�n le proporcionar� muchas otras herramientas para ayudarlo a ser el l�der de su salud y bienestar en general. Ademas de c�mo puede tomar el control de su propio bienestar. Incluyendo el trabajo de laboratorio que debe solicitar a su m�dico y c�mo entender qu� significa la informaci�n y qu� debe hacer con ella. Una de las formas m�s comunes en que los m�dicos utilizan los laboratorios es evaluar qu� es lo que no funciona cuando el paciente no se siente bien y analizar el final de un continuo de la enfermedad. Si su funci�n cambia, sus c�lulas hep�ticas pueden estar muriendo. Sin embargo, si son normales, muchos m�dicos dicen: “Oh, usted es normal”. Pero en realidad puede no ser normal. La buena noticia es que puede encontrar desequilibrios antes para tratarlos a tiempo.
Aunque est� cambiando lentamente, muchos m�dicos tienen la costumbre de no darles a los pacientes sus an�lisis de laboratorio reales. Y si este no es el caso, varios m�dicos no brindan explicaciones detalladas de las pruebas de laboratorio del paciente que no sean “Sus pruebas de laboratorio regresaron normales” o “Su colesterol esta un poco alto” o “Su nivel de az�car en la sangre esta alto”. Como quiropr�ctico, creo que todos deber�an tener acceso a sus an�lisis de laboratorio y que estos deben explicarsele al paciente. Tenemos que empezar a democratizar la medicina. Y esto se ha vuelto m�s fundamental que nunca.
Ahora, �por qu� es esto tan importante? Aproximadamente 133 millones de estadounidenses se ven afectados por enfermedades cr�nicas y ese n�mero es a�n mayor dependiendo de c�mo se define la enfermedad cr�nica. Aproximadamente uno de cada dos individuos en los Estados Unidos tiene pre-diabetes o diabetes tipo 2. La tasa de una variedad de enfermedades, incluyendo problemas digestivos, enfermedades al�rgicas, enfermedades del coraz�n, enfermedades autoinmunes, c�ncer, obesidad, diabetes tipo 2 y demencia, ha aumentado. Donde aproximadamente uno de cada tres ni�os nacidos hoy tendr�n diabetes tipo 2 durante su vida y una de cada dos personas mayores de 85 a�os y una de cada cuatro personas mayores de 75 a�os tendr� demencia.
Problemas de salud como estos se est�n manifestando en toda nuestra poblaci�n a un ritmo tremendo y pueden afectar a todos, ya sea personalmente o a trav�s del sufrimiento de un ser querido. Adem�s, las enfermedades cr�nicas han causado una carga econ�mica dram�tica en nuestro pa�s.
Sin embargo, la raz�n por la que estoy tan feliz de compartir este video con ustedes es porque hay mucho que podemos hacer para cambiar el futuro de nuestro sistema de atenci�n m�dica. Muchos m�dicos y yo tenemos el conocimiento que necesitamos para disminuir o incluso eliminar el sufrimiento de tantas personas y para salvar la econom�a. Todos los d�as en mi consultorio, veo que los pacientes recuperan su calidad de vida despu�s de que esperaban sufrir toda su vida debido a enfermedades cr�nicas como el reflujo �cido, el s�ndrome del intestino irritable, los dolores de cabeza, la fatiga y la artritis, entre otros problemas de salud. Problemas como alergias, problemas hormonales, obesidad, diabetes, enfermedades del coraz�n, enfermedades autoinmunes y depresi�n. Y los pacientes realmente pueden mejorar y prosperar, no solo enfrentar o controlar su enfermedad. En unos pocos meses, incluso semanas, de visitarnos a m� y a mi personal, las vidas de los pacientes pueden cambiar enormemente.
La medicina funcional analiza c�mo funciona el cuerpo humano por completo, y su enfoque m�s b�sico es comprender primero los factores, los genes y los desencadenantes de la enfermedad, y c�mo el estilo de vida y las aportaciones ambientales, incluyendo la dieta, el estr�s, las toxinas, los al�rgenos y los microbios , interact�an con el cuerpo humano para crear desequilibrios que com�nmente pueden conducir a problemas cr�nicos de salud.
Entonces, d�jenme tomar un momento para discutir algo. El cuerpo humano es un sistema. Y todo este sistema est� formado por “sistemas peque�os” que interact�an din�micamente. Pero, cuando uno o m�s de estos sistemas se desequilibran, puedes enfermarte. Y cuando estos sistemas se equilibran de nuevo, te vuelves sano. Y eso es lo que es la medicina funcional. La medicina funcional es simplemente comprender las causas de los desequilibrios en el cuerpo humano y el tratamiento para restablecer el equilibrio y proporcionar necesidades esenciales a todos los sistemas del cuerpo humano. Creando salud y bienestar en general es una ciencia. Y los m�dicos logran esto utilizando el historial m�dico detallado de un paciente, combinado con pruebas de laboratorio espec�ficas. Los m�dicos que siguen un enfoque de medicina funcional, como yo, a modo de ejemplo, generalmente eval�an su intestino y su microbioma, que no son muchos los m�dicos que analizan esto. Tambi�n queremos analizar su sistema inmunol�gico y si la inflamaci�n lo est� afectando, algo que llamamos defensa y reparaci�n. Y queremos saber c�mo sus mitocondrias producen energ�a con los alimentos y el ox�geno. Los profesionales de la medicina funcional desean saber si tiene alguna disfunci�n en la producci�n de energ�a, que suele ser la fuente de numerosas enfermedades, como la enfermedad de Alzheimer y el autismo, la diabetes tipo 2 y la fatiga, entre muchas otras. Tambi�n determinamos su carga t�xica y su capacidad de desintoxicaci�n, involucrando la funci�n de otros sistemas de comunicaci�n en su cuerpo, como sus hormonas. Finalmente, evaluamos su sistema estructural, desde sus c�lulas hasta sus estructuras biomec�nicas y c�mo �stas interact�an con sus creencias, emociones y m�s.
He usado estos procedimientos, una y otra vez, para ayudar a revertir las enfermedades cr�nicas en mis pacientes y educarlos sobre c�mo lograr la salud y el bienestar a largo plazo, al mismo tiempo que aprended como convertirse en las mejores versiones de s� mismos. Todos tenemos la oportunidad de curar o mejorar enormemente los problemas de salud o problemas que a menudo se diagnostican con la medicina convencional. La medicina funcional ofrece la oportunidad de descubrir el bienestar general a cualquier edad.
De hecho, trat� a un hombre llamado George, que ten�a 63 a�os, pesaba 300 libras y ten�a una variedad de problemas de salud. Ten�a reflujo �cido, problemas de sinusitis, diabetes, problemas card�acos, angina, problemas de pr�stata, disfunci�n sexual y edema en las piernas. Y como resultado, estaba bajo una amplia gama de medicamentos. Un d�a vino a mi oficina y me dijo: “�Me puede ayudar?”. Le dije: “S�, pero tendr� que hacer todo lo que yo le diga”.
Entonces, le recomend� una dieta anti-inflamatoria, que consist�a de alimentos integrales y una cantidad baja de az�car, y le aconsej� que tomara una variedad de suplementos para optimizar su nutrici�n, mientras lo guiaba a participar en ejercicios y actividades f�sicas. Dentro de un a�o, fue como un milagro, el paciente hab�a perdido 155 libras, revirti� todos sus problemas de salud y dej� de usar sus medicamentos. Ahora, �l est� planeando el resto de su vida, cuando antes de venir a visitarme, estaba planeando el final de su vida. Otra paciente que trat�, llamada Isabel, luch� contra una enfermedad autoinmune. Ten�a solo 10 a�os y ya estaba bajo un mont�n de drogas, esteroides, inmunosupresores e incluso quimio. Resulta que su dieta y la exposici�n a metales pesados ??estaban afectando sus intestinos y causando inflamaci�n. Ahora, simplemente trat� la fuente de sus problemas y su enfermedad autoinmune desapareci�. Dej� de usar sus medicamentos recetados y, en conjunto, su calidad de vida mejor� y prosper�.
La medicina funcional es un enfoque de tratamiento alternativo que se centra en las interacciones entre factores externos o ambientales, as� como factores internos asociados con los sistemas gastrointestinal, endocrino e inmunol�gico del cuerpo humano. Encontrar el m�dico adecuado puede hacer una gran diferencia cuando se trata de obtener atenci�n m�dica en vez de atenci�n por enfermedad. La medicina funcional trata la fuente del problema en lugar de tratar solo los s�ntomas. Como quiropr�ctico y practicante de medicina funcional, mi objetivo principal es brindar a los pacientes la atenci�n que merecen por sus problemas de salud, as� como educarlos sobre los fundamentos de la medicina funcional. El prop�sito de este art�culo es ayudar a los pacientes a encontrar al m�dico adecuado y establecer una relaci�n sana entre m�dico y paciente. Dr. Alex Jimenez D.C., C.C.S.T.
Informacion General Sobre el el Cuidado de Enfermos y el Cuidado de Salud
Durante las pr�ximas semanas, repasaremos algunos de los principios y conceptos fundamentales que me ayudaron a encontrar la causa subyacente de la enfermedad a trav�s del uso de la medicina funcional. Y eso los ayudar� a alcanzar la salud y el bienestar general.
En nuestro pr�ximo video, le ense�ar� c�mo puede tomar sus propias medidas para comprender su propio bienestar y qu� puede hacer para mejorar sus signos vitales. Este procedimiento ayudar� a proporcionarle datos e informaci�n de referencia importantes para ayudarlo a comprender mejor sus riesgos de salud. Tambi�n le permitir� evaluar y analizar su propio progreso a medida que trabaja para alcanzar sus objetivos de bienestar finales.
En el tercer video, voy a tratar de cubrir todo sobre nutrici�n. Le explicar� c�mo podemos evaluar su estado nutricional y c�mo puede utilizar los alimentos como tratamiento para comenzar a cambiar su salud y bienestar general lo mas antes posible. Su dieta es una de las partes m�s eficientes que puede controlar para crear un estilo de vida m�s saludable. Estar� compartiendo consejos y trucos para una mejor nutrici�n.
Debido a que las hormonas pueden afectar casi todos los aspectos de nuestra salud, tambi�n los vamos a destacar en el cuarto video. Desafortunadamente, la mayor�a de los m�dicos no entienden los niveles �ptimos de hormonas ni son conscientes de los m�todos adecuados para evaluar las hormonas. Lo preparar� para tener una conversaci�n efectiva con su m�dico sobre qu� tipo de prueba hormonal realmente importa, qu� significan y qu� puede hacer al respecto.
En el video cinco, me centrar� en la salud del coraz�n, discutiendo la presi�n arterial alta, colesterol alto y enfermedades cardiovasculares. Estas condiciones son extremadamente comunes, y desafortunadamente, la medicina convencional frecuentemente trata de tratarlas simplemente controlando los s�ntomas. Reducir el colesterol, disminuir la presi�n arterial, eso no resuelve la ra�z de los problemas. La mayor�a de las veces, el aumento de los niveles de az�car en la sangre, la obesidad y la diabetes son la causa de las enfermedades cardiovasculares. Voy a demostrar c�mo podemos prevenir y revertir estas enfermedades utilizando modificaciones en su estilo de vida, incluyendo la nutrici�n, para mejorar la salud del coraz�n.
En el sexto video, hablar� sobre la obesidad y la diabetes, trastornos metab�licos que van desde la grasa m�nima y la resistencia moderada a la insulina hasta la prediabetes y la diabetes tipo 2. Lo bueno de estos trastornos metab�licos, sin embargo, es que son completamente reversibles. Pero la mayor�a de los m�dicos convencionales no saben que es reversible. Voy a ense�arles c�mo pueden recuperar un metabolismo saludable y c�mo revertir la gama de problemas relacionados con la diabetes, que, como aprender�, son muchas enfermedades diferentes.
En el video siete, hablar� sobre el sistema inmunol�gico, en particular sobre la inflamaci�n oculta y la enfermedad. Como se describe que la inflamaci�n es la causa subyacente de la mayor�a de las enfermedades cr�nicas, continuamos observando tasas crecientes de enfermedades autoinmunes y alergias, que son se�ales de que nuestro sistema inmunol�gico no funciona correctamente. Sin embargo, la inflamaci�n tambi�n est� asociada con la obesidad, la diabetes tipo 2, las enfermedades del coraz�n, el c�ncer, la demencia e incluso la depresi�n. Le explicar� c�mo puede hablar con su m�dico sobre las pruebas de inflamaci�n, qu� significan los marcadores y c�mo puede cuidar su sistema inmunol�gico a un nivel m�s profundo.
En el video ocho, vamos a pasar al tema del metabolismo y las mitocondrias. Ahora, cada una de nuestras c�lulas contiene cientos o miles de mitocondrias, los generadores de energ�a de nuestras c�lulas. Cuando estos se da�an, podemos sufrir una variedad de problemas, como dolor, p�rdida de memoria, fatiga y muchos otros s�ntomas. Voy a explicar c�mo podemos cuidar sus mitocondrias a nivel celular y por qu� esta es una pieza vital de salud y bienestar �ptimos.
Luego, en el video nueve, vamos a explorar el significado de la desintoxicaci�n o el sistema de desintoxicaci�n del cuerpo humano y por qu� esto es una parte esencial del proceso de curaci�n. Desde el moho hasta los metales pesados ??y otras toxinas, explicar� lo que debe saber para evitar las toxinas y c�mo puede eliminar las que ya tiene para optimizar su capacidad de desintoxicaci�n. Con la cantidad de toxinas a las que estamos expuestos de manera regular, es importante identificar las causas que pueden estar afect�ndote y c�mo puedes comenzar a eliminarlas de inmediato. Afortunadamente, esto es algo que puede hacer f�cilmente para crear un estilo de vida m�s limpio para usted y sus seres queridos.
Finalmente, en el video diez estaremos hablando sobre la digesti�n. La salud digestiva es uno de los temas m�s discutidos en la medicina funcional. Debido a que nuestro sistema digestivo es el centro de nuestra salud, al diagnosticar problemas en este sistema, podemos mejorar todo, desde la salud mental, la absorci�n de nutrientes y el riesgo cardiovascular hasta la funci�n del sistema inmunol�gico. En el d�cimo video, los guiar� a trav�s de las pruebas disponibles para controlar su salud intestinal, as� como lo que puede hacer para mejorar su propia salud digestiva, lo antes posible.
Estoy muy contento de que me acompa�en en esta serie de videos porque el futuro de la medicina depende de ello. Cuando se aprende c�mo puede impactar su bienestar al ser proactivo y al asociarse con su m�dico, todo cambia. A trav�s de la medicina funcional, tambi�n puede ayudar a transformar nuestro sistema de atenci�n m�dica en un sistema de atenci�n m�dica real.
Entonces, terminemos este video con las preguntas que desea hacerle a su m�dico cuando los entreviste para ver si pueden ser un socio para usted y su salud y bienestar en general. Estas son solo algunas preguntas de alto nivel para comenzar con su discusi�n. �Est� dispuesto a trabajar conmigo como socio para mi bienestar?, �Cu�l es su punto de vista sobre nutrici�n y salud? �Cree que la comida es medicina? �Est� dispuesto a darme copias de los resultados de mis pruebas y explicarme qu� significan? La raz�n por la que nos hacemos estas preguntas es para asegurarnos de que su m�dico est� dispuesto a trabajar con usted y comprender el papel de la medicina funcional.
�
Estoy muy feliz de ser parte de su camino hacia una mejor salud y bienestar. Muchas gracias por acompa�arme el dia de hoy. El alcance de nuestra informaci�n se limita a problemas quiropr�cticos y de salud de la columna, as� como a temas y discusiones de medicina funcional. Para seguir discutiendo el tema, no dude en preguntarle al Dr. Alex Jimenez o comun�quese con nosotros al�915-850-0900�.
Curado por el Dr. Alex Jim�nez
Discusi�n del Tema Adicional: Dolor de Espalda Agudo
El dolor de espalda es una de las causas m�s frecuentes de discapacidad y d�as perdidos en el trabajo en todo el mundo. El dolor de espalda se atribuye a la segunda raz�n m�s com�n para las visitas al consultorio del m�dico, superada �nicamente por infecciones respiratorias superiores. Aproximadamente el 80 por ciento de la poblaci�n experimentar� dolor de espalda al menos una vez a lo largo de su vida. La columna vertebral es una estructura compleja formada por huesos, articulaciones, ligamentos y m�sculos, entre otros tejidos blandos. Las lesiones y / o afecciones agravadas, como las hernias de disco, pueden provocar s�ntomas de dolor de espalda. Las lesiones deportivas o las lesiones por accidentes automovil�sticos suelen ser la causa m�s frecuente de dolor de espalda; sin embargo, a veces los movimientos m�s simples pueden tener resultados dolorosos. Afortunadamente, las opciones de tratamiento alternativo, como la atenci�n quiropr�ctica, pueden ayudar a aliviar el dolor de espalda mediante el uso de ajustes de la columna vertebral y manipulaciones manuales, lo que finalmente mejora el alivio del dolor. �
De XYMOGEN�Las f�rmulas profesionales exclusivas est�n disponibles a trav�s de profesionales de atenci�n m�dica con licencia seleccionados. La venta por internet y el descuento de f�rmulas XYMOGEN est�n estrictamente prohibidos.
Con orgullo�El Dr. Alexander Jimenez�hace que las f�rmulas de XYMOGEN est�n disponibles solo para los pacientes bajo nuestro cuidado.
Llame a nuestro consultorio para que podamos asignar una consulta m�dica para acceso inmediato.
Si eres paciente de�Injury Medical & Chiropractic�Clinic, puedes preguntar por XYMOGEN llamando�915-850-0900.
Para su conveniencia y revisi�n de la�XYMOGEN�productos por favor revise el siguiente enlace. *XYMOGEN-Descargar-Catalogo
* Todas las pol�ticas de XYMOGEN anteriores se mantienen estrictamente.
Each chemical reaction which occurs in the human body requires enzymes and each one of these processes needs a coenzyme. But what are coenzymes? They are vitamins and minerals. Approximately 37 billion, billion chemical reactions occur in the human body every second.
That is why proper nutrition and a balanced diet rich in whole foods with vitamins and minerals is fundamental towards overall health and wellness. The majority of people in the United States are vitamin and/or mineral deficient. But, how do you know if you�re a part of the 90 percent of individuals with enough deficiencies to develop disease? We will discuss the tests you can utilize to find out if you�re vitamin and/or mineral deficient and what you can do about it.
What�is�Nutrition?
Hello, welcome to part three of �Taking Control of your Healthcare�. Today, we will discuss one of the fun topics of functional medicine: nutrition. Unfortunately, nutrition is one of the most essential conversations that many doctors aren�t willing to have with their patients. The average medical doctor learns about disease and malnutrition rather than learning how to use nutrition as treatment or even how to use nutritional therapies to achieve optimal health and wellness.
I personally believe that food can be utilized as a form of medicine. That it should be the foundation of medical practice, not an afterthought in medicine. There is no better treatment than proper nutrition. Approximately 90 percent of individuals in the United States aren�t getting the essential nutrients they require for healthy bodily functions. And more than that probably aren�t getting enough nutrients to prevent diseases associated with nutritional deficiencies. However, what is ultimately needed to achieve optimal well-being? More than 98 percent of Americans are deficient in omega-3, 80 percent in vitamin D, 50 percent in magnesium, and 10 percent in vitamin C. Nutrient deficiencies can also continue to cause health issues for years.
Acute diseases, such as rickets, scurvy, beriberi, or iron deficiency anemia, are often the most talked about health issues associated with nutrient deficiency, however, there�s also something known as long latency deficiency diseases. So, how much vitamin D do we need to not get rickets? Not a lot, only 30 units really. And how much do we need to not get osteoporosis? Perhaps about 3,000 to 4,000 units per day. Now, how much folate do we need to not get anemia? Also not very much. But, how much do we need to prevent heart disease, cancer, and dementia? You definitely need a lot more units per day.
Each chemical reaction which occurs in the human body requires enzymes and each one of these processes needs a coenzyme. But what are coenzymes? They are vitamins and minerals. Approximately 37 billion, billion chemical reactions occur in the human body every second.
That is why proper nutrition and a balanced diet rich in whole foods with vitamins and minerals is fundamental towards overall health and wellness. The majority of people in the United States are vitamin and/or mineral deficient. But, how do you know if you�re a part of the 90 percent of individuals with enough deficiencies to develop disease? There are only several nutrients which we are generally tested for. And for a majority of these, doctors aren�t aware of what the optimal values should be which can make correcting the nutrient deficiency so much difficult to do.
Taking Control of Your Nutrition
One of the most fundamental nutrients you need to measure is vitamin D. Although it�s referred to as a vitamin, it�s actually more like a hormone and it�s produced from cholesterol. This is yet another reason why cholesterol is essential. Approximately 80 percent of the population is deficient in vitamin D. Unless you�re in the sun 20 minutes every day between 10:00am and 2:00pm, you might need to take vitamin D supplements. In order to supplement properly, however, we need to know from what level you are starting at first. By way of instance, optimal vitamin D levels should be anywhere between 50 and 80 nanograms per milliliter of blood. The recommended amount of vitamin D we can supplement is about 2,000 to 4,000 units.
If you have lower vitamin D levels or if you have genetic problems, you may actually need to supplement with up to 10,000 units of vitamin D. That�s why it�s fundamental to work with a doctor or functional medicine practitioner who can measure and test your nutrient levels as well as help you optimize them. Most supplements contain about 400 units which is 10 times less than the amount most of us need. The optimal levels are generally just over 20. This is way too low. In one research study, women with vitamin D levels between 45 and 60 experienced reduced preterm labors by up to 60 percent. Vitamin D is also essential to help build strong bones and muscles, to improve immune system function, to prevent cancer, and ultimately, to help you live longer. It�s incredible.
Another measurement or test that�s performed by most doctors but is not always interpreted correctly is referred to as the MCV or mean corpuscular volume. The MCV measurement evaluates the size of your red blood cells in a test called CBC, or complete blood count, which is one of the most common blood panels ordered by healthcare professionals. So, if you are deficient in nutrients, your cells can either become smaller or larger. By way of instance, if your cells are too big, it could be a signs of a folate or vitamin B12 deficiency.
B vitamins are essential in numerous chemical reactions within the human body. They help us produce energy as well as help us regulate gene expression in order to create proteins that will ensure our overall health and wellness. If our B vitamins are too low, we could eventually develop an iron deficiency, anemia, or it could even cause a genetic disorder.
Optimal levels of B vitamins should be between 80 to 90. B complex vitamin supplements can help easily optimize levels of B vitamins. But, why would anyone be deficient in B vitamins? Is their diet not providing them with enough nutrients? Are they vegan? Are they taking any drugs and/or medications that prevent vitamin B12 absorption? Moreover, B vitamins are depleted during times of high stress which, as a practicing chiropractor, I can say it happens frequently to a majority of the population in the United States alone.
MCV is not the only measurement or test which evaluates a patient�s levels of B vitamins. Homocysteine is an alternative marker we will discuss in future articles which demonstrates B6, folate, and B12 levels. However, both the MCV and the homocysteine measurement or test only demonstrates that one or more of these nutrients may be deficient. It doesn�t necessarily tell us which one. Therefore, some additional, follow up evaluations may be required.
The MMA, or methylmalonic acid, measurement or test also shows vitamin B12 levels. Ultimately, vitamin B12 is essential for many processes in the human body, including energy production, gene expression, methylation, nerve function, and mood, among many other processes. Vegans have a higher chance of developing a B12 deficiency because it�s only found in animal products. Folate is another fundamental B vitamin. It can be determined directly in the blood, but, homocysteine is a more precise marker for folate levels.
In this section, we�re also going to discuss genetics because there is a measurement or test which can demonstrate a lot more regarding the status of your B vitamins and your ability to utilize them. Our genes are capable of making proteins. We have approximately 20,000 genes which are designed to create proteins. And one third of all the proteins they make are for our enzymes. Enzymes convert molecules into other molecules. These enzymes are also largely dependent on specific nutrients. One of the most fundamental genes which can be affected is known as MTHFR, or methylenetetrahydrofolate reductase. But you can just call it MTHFR.
MTHFR is essential because it helps regulate methylation, homocysteine, and folate, which are vital towards our overall health and wellness. When you have elevated levels of homocysteine, you should check your methylation status by looking for the MTHFR gene through a simple blood test.
Methylation is a key biochemical process which is fundamental towards the proper function of most of the human body�s systems. It triggers billions of times each second. And it ultimately helps control homocysteine, a substance which can damage blood vessels and has been associated with dementia, heart disease, and cancer, among other health issues. Methylation also helps repair your DNA on a regular basis as it helps recycle molecules necessary for detoxification, or getting rid of toxins. It also helps control your mood and it helps manage inflammation. Methylation is critical.
But, to make sure that methylation is active, the human body needs optimal levels of B vitamins. Without enough B vitamins, the methylation process can break down and the effects can be destructive. This is where we start seeing an increase in birth defects, such as spina bifida, down syndrome, and more miscarriages.
MTHFR is frequently abnormal in approximately 35 percent of the population. Methylation breakdown can also increase the risk of developing health issues like osteoporosis and diabetes, cervical dysplasia or cancer, including colon cancer and lung cancer, and even depression, pediatric cognitive dysfunction as well as mood and behavioral disorders, dementia, and stroke. Methylation is truly a key biochemical process.
When we discuss genetics, we have to understand that our environment can alter our genes. So, what if you have an MTHFR variation in your genes? First of all, not all mutations cause health issues. One mutation, by way of instance, known as C677T, is one version of the gene which is more significant than another version of the gene, known as A1298C. Now there�s no need to worry about these gene variations. They serve as examples to demonstrate you the quality of these mutations and how they function. People with these variations of the gene, by way of instance, might only need more folate or they might need a particular type of folate known as methylfolate. This is where a functional medicine practitioner can help their patients.
A genetic test can let you known if you have one of these gene variations. But, don�t get stressed. There�s a lot you can do to optimize your overall health and wellness. Many patients have visited my office after they find out they have these variations in their genes. And they quickly learn that they do have the option to take control of their well-being. However, what you do control is not your genes, you control your gene expression.
If you alter your healthy eating habits, you alter your nutrients. If you alter your environment, you alter which genes become active and which genes become inactive. And with these mutations, you can do just about the same thing by simply following the proper nutrition. When you find a doctor or functional medicine practitioner that�s willing to work with you, they�re going to tell you what lifestyle modifications you should follow to prevent health issues.
So, we�ve only just discussed the B vitamins. Next, we will discuss another fundamental nutrient in the human body: magnesium. Magnesium is a super essential mineral. Approximately 48 percent of people in the United States consume less than the required amount of magnesium from food. Magnesium is necessary in over 300 chemical reactions in the human body. It is also fundamental in the production of ATP, or the energy the human body utilizes as fuel.
A magnesium level blood measurement or test can help determine if you have a deficiency. Magnesium can also help reduce anxiety, calm the nervous system, and improve sleep. It is also an essential nutrient in the management of blood sugar levels. If you�ve been told by a healthcare professional that you have an average blood sugar level of over five and a half in something known as A1c, then magnesium can help.
Also, it�s very easy to know if you have a magnesium deficiency by looking at your current diet and symptoms. Do you eat enough magnesium rich foods like dark, leafy greens, beans, nuts and seeds? Or do you eat a lot of processed foods? Perhaps you also have symptoms such as anxiety, insomnia, constipation, muscle twitching, muscle cramps, PMS, and/or palpitations. If you have one or more of the symptoms I just mentioned, you may have a magnesium deficiency.
Next, we will talk about zinc, the immune-boosting and testosterone-boosting mineral in the human body. This important nutrient is in charge of maintaining your hair volume as well as repairing your gut lining. It�s also responsible for making sure your thyroid is functioning properly. Zinc can be easily measured or tested in the blood and unfortunately, it�s another nutrient we are highly deficient in, in the United States. Additionally, you can also look at your alkaline phosphatase levels, which can be calculated through a liver function evaluation on a regular blood panel. High levels of alkaline phosphatase may indicate the presence of cancer or bone problems, among other health issues, however, low levels of alkaline phosphatase may indicate a zinc deficiency, because it�s a zinc-dependent enzyme.
Finally, the last fundamental nutrient we are going to discuss is iron. Iron is frequently deficient in vegans and vegetarians, or in women in general due to menstruation. Iron is necessary for transporting oxygen throughout the human body and it�s ultimately essential for brain health and wellness. Iron is also important for hair and nails, sleep, and so many other things.
Ferritin is a stored type of iron and it�s this nutrient which helps you see your iron levels. Optimal ferritin levels should be between 50 to 150 in women and 100 to 300 in men. And many times I�ve seen women visit my office who have ferritin levels of less than 50, or worse, in the single digits. This is because pre-menopausal women lose blood every month due to their menstrual cycles and it becomes so much harder for them to maintain proper ferritin levels. Many women also eat way less than what they�re supposed to be eating every day. High levels of ferritin, on the other hand, could be a sign of inflammation, generally caused by insulin resistance to sugar, or it could be a sign of hemochromatosis or iron storage disease, a very dangerous genetic disorder.
Having decreased levels of ferritin can also make you feel tired, and it can cause hair loss, it can cause insomnia. So, even if your blood count is normal, if your ferritin levels are low or your iron levels are low, it can also cause these symptoms. That�s why if you experience symptoms of fatigue, it�s essential to measure or test your ferritin levels. And it can be easily supplemented.
Aside from ferritin, a low MCV can also determine if you have an iron deficiency. Iron deficiencies can cause red blood cells to become very small and that can be demonstrated in low MCV levels, which evaluate the size of your red blood cells. Additionally, transference saturation, serum iron, TIBC, or total iron binding capacity, and hemoglobin, can provide us with a more in depth look at your iron status to distinguish different causes of anemia. These are included on a regular iron blood panel in a lab test.
We�ve discussed several nutrients which can be ordered by a majority of healthcare professinals with access to conventional lab testing. Furthermore, there�s another test which can tell us more about which type of nutrients we need based on our genes. It�s called the DNA health test and it�s provided by a company called DNAlife. This test evaluates a variety of genetic markers associated with detoxification, lipid metabolism, and inflammation, including the MTHFR gene and other B vitamin markers. Now, DNA Health demonstrates the different genes we evaluate. And most of these are common genes, they�re those we can do something about. We analyze the genes we can change based on your nutrition and other lifestyle factors.
It shows us the MTHFR gene, other B vitamin markers, genes that control B6, folate, and B12 as well as demonstrating how they function and whether you have nutrient deficiencies. Then it tells us which nutrients you will need to supplement and how much we will need to give to you. It�s tremendously helpful.
There was an individual who had two variables of the MTHFR gene. This woman had miscarriage after miscarriage after miscarriage. She visited her doctor for an evaluation and it turns out that she had a folate-regulating mutation. So her doctor then started giving her the proper amount of folate she needed and she started having healthy babies. Sometimes, nutrition can be that powerful towards improving a patient�s overall health and wellness.
The DNA health test can help personalize your approach when optimizing your well-being based on your genetics. What we measure utilizing the DNA health test provide well-established insights about your genes as well as what you can do about them.
A micronutrient test known as the individualized optimized nutrition profile or the ION panel, are alternative test options which can also provide information about your current nutritional status. This test is by Genova. This is a robust test which measures all the essential vitamins and minerals, fatty acids, organic acids, and antioxidants you currently have. Ultimately, this test looks for imbalances, insufficiencies, or deficiencies, rather than looking for a specific disease. It looks for things that a majority of doctors never look at.
Functional medicine practitioners or doctors look at patient�s amino acid levels, mineral levels, and even toxin levels from heavy metals like mercury, lead, arsenic, and many more. We also look at your antioxidant levels, vitamin A and vitamin E levels, as well as your CoQ10 antioxidant and beta carotene status. We can determine if a person eats vegetables or not if, by way of instance, they have low levels of beta carotene. We also look at vitamin D levels, essential fatty acids, including your omega-3 fats and your omega-6 fats. We can tell if a person eats junk food. We can tell if a person is eating fish. And We can tell if a person is eating too much olive oil or saturated fats. It�s all demonstrated in these measurements and tests.
An OAT test, or organic acids test, also looks at what is known as organic acids. This test demonstrates a wide array of parameters associated with your mitochondria, which we will discuss in the next article, your B vitamins, your neurotransmitters, your gut flora, and your detoxification. It�s ultimately a comprehensive test which shows me if a patient is well or sick. It shows me where the imbalances are and where I need to recommend lifestyle modifications. It also helps provide clues about other health issues.
By way of instance, if your mitochondria aren�t functioning correctly because you have decreased levels of essential amino acids or you have increased oxidative stress or if you simply have low levels of selenium and zinc, there�s a possibility that you might have some form of toxic overload due to heavy metals. And that�s precisely what I would go looking for. Signs like these provide a lot of information about what we can do to treat a patient. And an experienced functional medicine practitioner or doctor can determine what�s really going on with a patient or they can help patients discover how to optimize their overall health and wellness.
Nutrition is the study of nutrients in food and how the human body utilizes nutrients as well as the relationship between diet, disease and overall health and wellness. Nutrients are a source of nourishment, including carbohydrates, proteins, fats, vitamins, minerals, fiber and water. Functional medicine focuses on the use of food as a form of medicine. A balanced nutrition can help prevent as well as treat a variety of health issues. Similarly, nutrition in functional medicine involves how certain diseases and conditions may be associated with dietary factors, such as poor diet or malnutrition, food allergies and food intolerances. Dr. Alex Jimenez D.C., C.C.S.T.
Understanding Your Nutrition
As good functional medicine doctors we�re often left asking ourselves, why is it that so many people in the United States are overfed but undernourished? Or, why is it that Americans eat too many calories and too few nutrients? The leading causes for the widespread nutritional deficiencies are the following: First, humans evolved from eating wild foods which contained tremendously higher levels of nutrients. Second, the soil we currently utilize to grow our crops in has become greatly depleted of nutrients. Hybridization techniques from industrial farming are yielding animals and vegetables to have decreased levels of nutrients. Third, processed foods have absolutely no nutrients, which is why they frequently have to be fortified. And last but not least, exposure to environmental toxins, lack of sunlight, chronic stress, and poor diet, including increased alcohol, caffeine, and sugar consumption, can increase our nutritional needs, much of which we�re already not getting enough from our current nutrition.
Well you might not need any vitamins, however, if you can meet certain conditions. Perhaps if you only hunted and gathered wild food and if you weren�t exposed to environmental toxins. Or maybe if you went to sleep with the sun and woke up with the sun, sleeping nine hours a night. And if you experienced absolutely no amount of chronic stress. Ultimately if you only drank pure, clean water and breathed pure, clean air. Then, you probably wouldn�t need any vitamins. But the rest of us that don�t follow these conditions, we do need them.
And with that thought, we wrap up this article. In the next article, we will talk about hormones. Hormones can affect almost every aspect of our well-being, and many healthcare professionals don�t understand what our optimal hormone levels should be or even when to test them and what to do about it once they do. Measuring and testing hormone levels should be standard practice, and many patients have never had a blood panel to look at their hormones. It�s fundamental to know as well as understand what�s going on inside your own body. And that�s why this next article is so important. You won�t want to miss our next update. See you soon.
The scope of our information is limited to chiropractic and spinal health issues as well as functional medicine topics and discussions. To further discuss the subject matter, please feel free to ask Dr. Alex Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topic Discussion:�Acute Back Pain
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief. �
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download
* All the above XYMOGEN policies remain strictly in force.
Today we will begin to discuss how the spectrum which medicine currently considers �normal� may not actually be optimal towards your overall health and wellness. These reference ranges can change based on age, gender, physical activity, and more. As a matter of fact, if we were to evaluate an individual�s weight in the United States, it would be considered �normal� to be overweight, simply because 70 percent of the population is overweight. Reference ranges for lab tests today are based on a sick population when we should aspire for optimal well-being.
Then, I will demonstrate how this knowledge can apply to the most basic medical measurements: your vital signs. Everyone knows that when you first visit a doctor, they take your vital signs, including your weight, your blood pressure, your heart rate, and your temperature. However, does your doctor tell you what your results mean? How can you tell if you�re healthy?
What�are�Vital�Signs?
Hello everyone, it�s Dr. Alex Jimenez. Welcome to part 2 of �Taking Control of Your Healthcare�. Today, we will discuss laboratory reference ranges associated with your vital signs. It may not sound like an interesting topic, but it�s really important for your well-being.
Most doctors generally define a patient�s test results as either �normal� or �abnormal�. But what exactly does �normal� and �abnormal� mean? Normal is when 95 percent of the population falls within a common range while abnormal is when the remaining percent of the population falls outside that common range. Whether you�re healthy or sick, young or old, although there can be some variations for children, normal and abnormal are simply statistical numbers which fall within two standard deviations.
However, these standard deviations don�t necessarily mean it�s optimal or not. It�s only a statistical number, after all. As a matter of fact, disease can occur along healthy and unhealthy individuals. These lab reference ranges should be defined according to what is best for a human being.
By way of instance, vitamin D levels are classified as normal if they�re over 20, however, the ideal levels are over 50. So why is 20 considered �normal�? This is because approximately 80 percent of the population is deficient in vitamin D, therefore, they fall under what is considered to be the �normal� range. However, this doesn�t mean that these levels are best for your overall health and wellness. Furthermore, �normal� blood sugar levels have been classified to be under 100, although we know �optimal� blood sugar levels have been classified to be between 70 to 80. Blood sugar levels over 80 can have an increased risk of disease. And unfortunately, our �normal� laboratory reference ranges are not optimal because we�ve become a sick population. In the United States it can be considered �normal� to be overweight because 70 percent of our population is overweight. But, although being overweight or obese is considered normal in the United States, that�s not something we would want to aspire to. We want to aspire to achieve overall health and wellness.
Now let�s continue to discuss the meaning of normal. Many patients often visit the doctor only to be told that their lab tests have returned normal, however, they may still be feeling sick. What does that mean? Does it mean you�re sick? Does it mean you�re healthy? As I�ve mentioned previously, either your doctor is missing something or you�re crazy, and I�m pretty sure your doctor is missing something. This is one of the main differences between conventional medicine and functional medicine. Through functional medicine, many doctors focus on health care rather than sick care. We�re looking for more subtle deviations from optimal.
Until your liver function is considered abnormal according to the current standard, your liver cells may already be dying. A functional medicine doctor may review your lab tests differently than a conventional doctor. This is primarily because the reference ranges that we�re focusing on aim towards optimal health, not disease. Many conventional doctors evaluate lab tests differently than functional medicine doctors, and then they either follow a �watch-and-wait approach�, or they label you as �not sick� after the most minimal amount of lab tests. As a matter of fact, one patient who visited me had blood sugar levels of 120, where a blood sugar of 126 is already considered type 2 diabetes. And I said, �Did you see your doctor regarding this?� And he said, �Yeah.� I asked him what the doctor said. And he finally said, �Well, he said to wait until I actually had diabetes and then to come back for medication.� And that is the last thing we want to be doing as healthcare professionals.
Reference ranges give us an average number of values which have been recorded among the general population. But let�s not forget, �normal� reference ranges are relative. They change based on age, gender, physical activity, and more. If you were to evaluate an individual�s weight in the United States today, it would be normal to be overweight, as I mentioned, only because 70 percent of people are overweight. And unfortunately, we keep changing our reference ranges based on our sick population. This is not what we should aspire to do as doctors. This is why functional medicine treats the individual, not only the numbers.
Additionally, reference ranges which were once considered normal can also change over time. One instance of how reference ranges change was demonstrated by a known global laboratory company called LabCorp, where they recently changed their reference ranges for male testosterone levels. Previously, LabCorp considered normal testosterone levels for an adult male to be between 348 to 1,197. This value was based on a population of lean adult males. However, in 2017, they lowered normal testosterone levels for an adult male to be between 264 to 916. Moreover, overweight men, excluding obese men, were probably included in the cohort study, ultimately changing reference ranges for male testosterone levels. Research studies have found that excess abdominal fat can cause lower testosterone levels. By changing reference ranges, however, this demonstrates that conventional medicine is considering overweight individuals to be a part of the norm. But this isn�t what we want. We want to strive for overall well-being.
This is why you need to start taking control of your own healthcare. As one in two people have some type of chronic disease, we have to evaluate how we interpret lab tests as well as how �normal� may not necessarily mean health and wellness but simply an average for a growing sick population in the United States.
Taking Control of Your Vital Signs
The primary goal of this series of videos is to encourage you to become the leader of your own well-being by understanding what your lab tests mean, understanding what optimal looks like, and understanding which lab tests are designed to help you achieve overall health and wellness rather than focusing on the disease. I would also like to educate you in order for you to make an informed decision on who you choose to be your doctor, or partner, in your journey to well-being. Now let�s look into the most common medical measurements: your vital signs.
Your vital signs are initially taken by the nurse when you visit the doctor. These vital signs generally include blood pressure, weight, heart rate, temperature, and even oxygen saturation. However, are you aware of what these numbers mean? Has your doctor discussed these numbers with you? Why would they take your vital signs if they�re just going to record them and never discuss them with you? Do the numbers actually demonstrate your health and wellness? If you have high blood pressure or a heart rhythm problem, your healthcare professional is most likely going to tell you, but otherwise, you may not find out what the value of your vital signs is.
Your heart rate is probably one of the most important vital signs taken during a doctor visit. The pulse is a measure of how fast your heart is beating. The human heart beats more than 115,000 times per day. So, if your heart rate is above 100, then we define that as having a high heart rate. But, if you have a heart rate higher than 80, that can increase your risk of developing heart disease. What causes this increase? Although many factors can lead to cardiovascular disease, stress is one of the most common causes because it raises your adrenaline and causes increased heart rate and blood pressure. Drinking too much coffee, stimulating medicines like Adderall, or an overactive thyroid, heart or lung health issue can also increase the risk of heart disease.
The fight-or-flight response, a physiological reaction which becomes activated during times of stress, can also cause an increase in heart rate. When an individual�s heart rate is frequently above 80, it might be time to begin incorporating some stress management techniques into their life, by way of instance, mindfulness meditation and other forms of meditation. Stress is not the only cause of an increased heart rate. Anxiety, magnesium deficiencies, being out of shape and dehydration can also cause an increased heart rate. Ideally, we want to achieve a lower heart rate, optimally under 70.
On the other side of the spectrum, a decreased heart rate below 60 might also demonstrate the presence of thyroid gland dysfunction or low thyroid function. Athletes and distance runners actually have low heart rates because they�re so well-conditioned. Their heart rates can be as low as 50 or even 45. But, if you have a low heart rate and you�re not an athlete or a distance runner, it may be time for you to go talk to your doctor.
While heart rate is one of the most important vital signs taken during a doctor visit, there�s another vital sign which can be just as important, your heart rate variability. This reflects the health of your automatic or autonomic nervous system, which is in charge of controlling all of the unconscious elements of your nervous system, such as digestion and breathing. Heart rate variability has frequently been associated with longevity and even death. The less variable the heart rate is, the higher the mortality rate. Many doctors don�t measure a patient�s heart rate variability, but fortunately, you can manage it yourself. Hot-and-cold therapies, saunas, exercise, yoga and meditation, can all help improve a patient�s heart rate variability.
Now let�s move on to the next most important of the vital signs taken during a doctor visit, blood pressure. If you were to line up all the blood vessels in your body, they would extend approximately 59,000 miles. That�s almost seven times around the earth. These same blood vessels carry over 7,500 liters of blood throughout your entire body on a regular basis. With each heartbeat, blood is pushed against the artery walls, which causes an increase in pressure.
Medical measurements for blood pressure have two numbers. The top number is known as systolic, or the pressure when the heart is contracting, and the bottom number is known as diastolic, or the pressure when the heart is relaxing or at rest. Normal reference ranges for blood pressure continue to change because we keep finding out that the reference ranges we used to think of as normal, which were first 140/90, then 130/80, were still associated with an increased risk of stroke and heart attack. Many doctors today may mention a problem with your blood pressure only if it�s over 130/80.
The reason why blood pressure is so important is because when it�s elevated, it can place additional pressure on the heart and arteries, potentially leading to heart disease, or stroke, heart failure, or even kidney failure. And it�s not only the heart and arteries which are affected by blood pressure: the brain, kidneys and even the eyes can all be tremendously affected, leading to strokes, dementia, kidney failure, and blindness, among other health issues. Maintaining your blood pressure at an optimal level is fundamental towards your overall health and wellness. As a matter of fact, normal blood pressure is currently believed to be under 120/80, however, it may turn out to be even lower.
While high blood pressure is bad, low blood pressure can also be just as bad. A good functional medicine doctor will discuss with you the risks of both high blood pressure and low blood pressure. Blood pressure below 100/60 may cause problems but not necessarily. Blood vessels in the human body function just like pistons in a car. If not enough pressure is built up, it can become really difficult for the blood to flow against gravity. And because the human brain is positioned higher than the heart, we depend on our blood pressure to supply our brain with the necessary amount of oxygen and nutrients required to function accordingly.
If you have low blood pressure, you might experience other symptoms, such as fatigue. Other symptoms associated with low blood pressure include dizziness when standing, weakness and even brain fog. Also, both chronic high blood pressure and chronic low blood pressure may contribute to an increased risk of dementia.
Now we�ve discussed the importance of heart rate and blood pressure. But, how about we discuss another important vital sign: your body temperature? A fever or an elevated body temperature can often be a sign of infection. Temperature can also provide an insight into the function of our metabolism. The lower an individual�s metabolism, the less heat they produce, which may manifest as a slightly lower-than-normal body temperature. The thyroid gland plays a big role in metabolism and regulating your temperature. So, if you frequently feel cold, you might want to discuss ordering the right thyroid panel with your doctor. But, what tests should you ask your doctor to determine this? Don�t worry, we will discuss which thyroid tests you should take in the video on hormones. The optimal body temperature should be approximately 98.6 degrees Fahrenheit. If it�s lower than 97.7 degrees Fahrenheit, however, it may indicate that you have a thyroid problem.
The final medical measurements we�re going to discuss are your height and your weight. Doctors utilize your height and weight to calculate your body mass index, or BMI. At our clinic, by way of instance, we utilize the InBody 770, a body composition and body water analyzer, to help easily determine your body mass index and more. However, body mass index doesn�t always factor in body composition, or the percentage of fat versus muscle in the human body. By way of instance, a pro football player who is 6�6� and 265 pounds has a body mass index of over 30, which puts them in the obese category.
But if you were to take a look at this individual�s body, they would never be categorized as obese. This demonstrates that BMI isn�t an accurate measurement, especially for athletes. Also, a 65-year-old woman may have more fat than muscle in their body while their BMI measurements may appear �optimal�. Instead, many functional medicine doctors use waist-to-hip medical measurements. This is a simple measurement you can do at home to determine body fat distribution, which can also help demonstrate the risk of metabolic dysfunction. Obesity, type 2 diabetes, and heart disease are caused by excess belly fat or fat accumulated around the organs. Excess fat around the midsection can ultimately increase the risk for heart disease and metabolic issues, such as diabetes, dementia, cancer and many other health problems.
But first, let�s discuss how you can calculate your waist-to-hip ratio. To measure your waist, you simply take the measurements of the widest area around your waist, which is generally the biggest part around your belly button. To measure your hip, you then take the measurements of the widest area around your hip, which is generally where your hip bones are on your sides. So, you take these measurements and then you divide the measurements of your waist by the measurements of your hip. And this is the most fundamental number you have to look at.
In men, a waist-to-hip ratio of less than 0.9 is considered optimal. If the ratio is greater than one, meaning that your belly is bigger than your hip, it can put men at higher risk of developing metabolic syndrome, heart disease, diabetes, stroke, cancer, and dementia. In women, a waist-to-hip ratio of less than 0.8 is considered optimal. If the ratio is greater than 0.85, it can put women at higher risk of developing metabolic syndrome as well as the other health issues mentioned above.
Medical measurements, including heart rate, temperature, respiration rate, and blood pressure, are several vital signs which help indicate doctors the state of a patient’s fundamental body functions. Reference ranges today are utilized to determine “normal” health and wellness spectrums, however, research studies have demonstrated that these reference ranges may actually not be optimal spectrums. Understanding the most basic medical measurements, or vital signs, is important towards a patient’s well-being, as it can help people recognize whether they are feeling healthy or sick, regardless of the standards.
Understanding Your Vital Signs
And those were your vital signs, your most basic medical measurements. These numbers are very essential as they are fundamental towards your overall health and wellness. Understanding how your body functions as a whole is important to optimize your well-being. Therefore, next time you visit your doctor, ask about your vital signs and discuss these reference ranges with them. I truly believe that a combination of your own research and having a good relationship with a qualified healthcare professional can lead you on the right path to overall health and wellness.
Finding a doctor that will work with you is essential towards achieving the results you deserve. If your doctor is not willing to have a conversation with you about your well-being or the lab tests that are needed to track your results, then you might want to consider finding another doctor.
If you learned about the simplest medical measurements then you will definitely enjoy the next video, where we will discuss the blood tests utilized to determine nutritional deficiencies. Over 90 percent of individuals in the United States are deficient in nutrients at the RDA level. That�s the minimum amount necessary to prevent diseases caused by nutritional deficiencies, such as scurvy and rickets.
We�re going to discuss how a doctor who specializes in functional medicine evaluates results and what other tests your doctor might be unaware of that can tell us a lot about your nutritional status. Thanks again for joining me so far and I�ll see you later. The scope of our information is limited to chiropractic and spinal health issues as well as functional medicine topics and discussions. To further discuss the subject matter, please feel free to ask Dr. Alex Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topic Discussion:�Acute Back Pain
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief.
XYMOGEN�s Exclusive Professional Formulas are available through select licensed health care professionals. The internet sale and discounting of XYMOGEN formulas are strictly prohibited.
Proudly,�Dr. Alexander Jimenez makes XYMOGEN formulas available only to patients under our care.
Please call our office in order for us to assign a doctor consultation for immediate access.
If you are a patient of Injury Medical & Chiropractic�Clinic, you may inquire about XYMOGEN by calling 915-850-0900.
For your convenience and review of the XYMOGEN products please review the following link.*XYMOGEN-Catalog-Download
* All the above XYMOGEN policies remain strictly in force.
Today we will discuss the fundamentals of functional medicine and how you can build a healthy doctor-patient relationship.
If you visit the doctor because you�ve been experiencing migraines, eczema, irritable bowel syndrome and depression, you�re most likely going to be referred to four different specialists and you may even be prescribed four different drugs and/or medications, at minimum. A functional medicine approach understands that there may be common underlying health issues which can be causing a patient�s symptoms. Once you get to the source of the problem, you can fix the health issues that create the symptoms.
Functional medicine asks, �Why do you have those symptoms and how can we treat the source of the problem and improve your overall health and wellness?� rather than, �What disease do you have and what drug do you use to treat it?�
What is Sick Care vs Health Care?
Hello everyone, my name is Dr. Alex Jimenez. I�m a chiropractor and practicing functional medicine doctor in El Paso, Texas. I�m so happy to introduce you to part one of �Taking Control of Your Healthcare�, where we will discuss the differences between �Sick Care and Health Care�. So, I frequently hear people talk about how difficult it is to find the right doctor, someone who is willing to work with them, who listens to them, and who is open to meeting their demands while teaching them everything they need to know about their problems. A doctor who accepts innovative advances in science and system approaches to determine the source of the problem. This is known as functional medicine, and we will discuss more of that in a minute.
My goal right now is to show you how you can find the right doctor and build the best relationship with them to get the care you deserve. It�s important to ask your doctor several questions to determine if they are the appropriate fit for you, if they�re willing to work with you, to listen to you, and if they�re open to your requests as well as learning about problems they�re not familiar with and following new treatment approaches. By way of instance, if the doctor you choose says that nutrition has nothing to do with disease, then you should probably go find another doctor. Now let�s discuss what questions you should ask your doctor regarding your overall health and wellness in order to be informed about what�s happening in your own body.
I�ve talked to patients who have visited numerous doctors in hopes of finding an answer as to why they�re not feeling well or have some type of disease. Many are frequently referred to one or more specialists and, in some cases, they�re given Prozac, or they�re told that their symptoms are all in their head or sometimes they�re even told that there�s nothing wrong with them after their lab results are all normal. And it�s frequently these same patients who are often sent to various doctors for each part of their body rather than being sent to a doctor who can diagnose and treat their body as a whole. If you go to the doctor and you have migraines, eczema, irritable bowel syndrome, and depression, in most cases, you�re going to be sent to four different specialists and you�re going to be given, at least, four different medicines instead of visiting a specialist who can understand the underlying source of the problem and treat the root of the symptoms.
Have you ever visited a doctor who, after explaining your symptoms to them, said, �Oh, I did a full blood panel and everything came back normal.� This could mean one of two things, either you�re crazy or they�re missing something. And I�m sure they�re often missing something because they�re not looking in the right places for the answers. It�s like the guy who dropped his keys in the street and when his friend sees him looking under a lamppost, he says, �Hey, what are you doing?� He says, �Well, I�m looking for my keys.� �So, where did you lose them.� He says, �Well I lost them down the street.� �Then why are you looking here?� �Well, the light�s better here.� And unfortunately, that happens a lot in medicine. Doctors will look for answers where problems are easy to find. And that is the purpose of this article. To help you understand how you can find the source of the underlying health issues that are making you feel sick.
This reminds me of a case of a woman I saw in my office who had psoriatic arthritis, an autoimmune condition which caused her to have rashes all over her body along with joint pain and swelling. As a result, she was on large amounts of drugs and/or medications which added up to about $60,000 dollars a year or more. She also had other health issues, she had acid reflux, irritable bowel syndrome, pre-diabetes, migraines, insomnia and depression. This woman visited many different specialists and she was taking prescriptions to address each of her symptoms. When it came time for me to see her, all I did was get to the source of the problem.
So, during diagnosis, I said to myself, �Okay, this patient has inflammation, but what�s the cause of her inflammation?� And instead of referring her to numerous other specialists so she could receive medicine for her migraines, acid reflux, depression, etc., I said to myself, �Oh, all of these symptoms are inflammatory, so what�s the root of the inflammation?� Well it turns out that the patient had problems with her gut all along. I then helped her clear out the �bad bugs� in her gut by recommending her an anti-inflammatory diet, I included some supplements, fish oil, vitamin D, probiotics. Honestly, really basic natural remedies. When she returned six weeks later, all of her symptoms had disappeared. She had gotten off all of her prescription medicines and she had lost 20 pounds. I didn�t tell her to stop taking her meds, she just did it on her own. It was absolutely remarkable and that�s what happens when you treat the source of a patient�s underlying health issues. And you don�t have to do a lot to get to the root of the cause.
I understand that it may not always be possible to work with a doctor who�s trained in functional medicine, which we will discuss what that means in a minute, however, it is possible to find a doctor who�s going to be willing to work with you, who�s going to listen, who�s going to have an open mind and who�s going to be your partner during your journey to overall health and wellness. Throughout this article, I�m going to discuss conventional lab testing as well as innovative functional medicine lab testing, demonstrating it all for the purpose of creating well-being rather than viewing it all from the perspective of the disease in order to find the source of a patient�s health issues and correct imbalances through a functional medicine approach.
Taking Control of Your Health Care vs Sick Care
Functional medicine is often referred to as the future of medicine, but it�s currently being offered by many doctors if you visit the right place. The purpose of functional medicine is to diagnose and treat the root cause of a variety of diseases by evaluating the body as a whole, rather than by analyzing each collection of organs independently through separate specialists. Functional medicine treats the whole system, not just the symptoms. Doctors who practice functional medicine frequently ask themselves, �Why does the patient have these symptoms and how can I fix the root causes and improve their overall health and wellness?� rather than �What disease does the patient have and what drug and/or medication do they use to treat it?�
I�ve been a practicing chiropractor for over 25 years. And I�ve witness countless of miracles every day. My patients don�t simply find relief from their symptoms, they truly achieve overall well-being. Unfortunately, our current healthcare system is broken. Many doctors are tremendously affected due to time constraints set by insurance companies to pay bills, causing them to rush through their appointments. In turn, this continuous cycle can frequently end up leaving patients with unanswered questions followed by frustration. Our current healthcare system makes people feel powerless, often keeping them stuck in disease.
As a part of this healthcare system, it�s important that we understand that some diseases are not as easily treatable as a cold. Several diseases also don�t just develop randomly. The majority of diseases which exist today are related to your environment and how these external as well as internal factors alike interact with your genes and lifestyle to influence your health and wellness.
What we know today as the �conventional medicine� approach is typically referred to as the �name it, blame it, and tame it� game. First, the doctor will diagnose the patient and provide them with a label, by way of instance, they say a patient has depression. Now they have the name of the disease. And finally, the doctor will treat the disease with prescription drugs and/or medications. In the end, the patient is left taking an antidepressant. However, depression can be caused by a wide array of factors and the solution isn�t necessarily an antidepressant. Not only is this type of practice outdated, it�s actually considered to be quite unsafe in the long-run. And it�s often not providing patients with the results they need. People are being misdiagnosed and mistreated, frequently being left sick without really getting the proper care they deserve.
Conventional medicine can also be helpful at the very end stages of some diseases as well as for acute diseases. If you have an emergency or you�re feeling very sick, if you break a bone, or if you have a raging infection, conventional medicine, acute care, drugs and/or medications can be the right solution and we should be highly grateful for them. But this is not the approach we need to prevent and cure chronic diseases. If we actually understood how to care for our bodies the way we should, most of us wouldn�t be feeling as sick as we do. And many people will walk around feeling sick, but you don�t have to, it�s not normal. Many doctors now understand that a change is required to turn our entire sick care system into one that actually supports health care.
I would like to empower you to help transform the future of the medicine field by taking your well-being into your own hands. Throughout the next articles I�m going to share with you how you can find a doctor that takes consideration of your personal values and beliefs so you can achieve the results you�re looking for while in a safe and comfortable environment. You can learn how to be your own health advocate and become a true partner with your doctor. And there are many other doctors and functional medicine practitioners like me who are waiting to help.
Furthermore, I�m going to share with you how you can find the best doctor and I�m also going to provide you with many other tools to help you be the leader of your overall health and wellness. How you can take control and make the calls for your own well-being. Including what lab work you should ask your doctor for, and how to understand what the information means and what you should do with it. One of the most common ways doctors utilize labs is to evaluate what goes wrong when a patient isn�t feeling well and to analyze the end of a continuum of disease. If your liver function changes your liver cells may already be dying. If they�re normal, however, many doctors say, �Oh, you�re normal.� But it may actually not be normal. The good news is that you may be able to find imbalances sooner in order to treat them in time.
Although it�s slowly been changing, many doctors have the custom of not giving patients their actual lab tests. And if this isn�t the case, several doctors don�t provide detailed explanations of patient�s lab tests other than, �Your lab tests came back normal,� or �Your cholesterol is a little high,� or, �Your blood sugar is a little high.� As a chiropractor, I believe that everyone should have access to their lab tests and that these should be explained to the patient. We need to start democratizing health care. And this has become more fundamental than ever before.
Now why is this so important? Approximately 133 million Americans are affected by chronic disease and that number is even higher depending on how chronic disease is defined. About one in two individuals in the United States has pre-diabetes or type 2 diabetes. The rate of a variety of diseases, including digestive problems, allergic diseases, heart disease, autoimmune diseases, cancer, obesity, type 2 diabetes, and dementia have all been increasing. Where approximately one in three kids born today will have type 2 diabetes in their lifetime and one in two people over the age of 85 and one in four people over the age of 75, are going to have dementia.
Health issues like these are manifesting throughout our population at a tremendous rate and they can affect everyone, either personally or through the suffering of a loved one. Additionally, chronic diseases have caused a dramatic economic burden within our country.
However, the reason why I�m so happy to share this article with you is because there�s a lot we can do to change the future of our healthcare system. Many doctors and I have the knowledge we need to decrease or even eliminate the suffering of so many people and to save the economy. Every day in my office, I see patients recover their quality of life after they were expecting to suffer their entire lives due to chronic diseases such as, acid reflux, irritable bowel syndrome, headaches, fatigue, and arthritis, among other health issues. Problems like allergies, hormonal issues, obesity, diabetes, heart disease, autoimmune diseases, and depression. And patients can truly get better and thrive, not just cope with or manage their disease. Within a few months, even weeks, of visiting me and my staff, the lives of patients can tremendously change.
Functional medicine looks at how the human body functions as a whole, and its most basic approach is to first understand the factors, genes, and triggers for disease, and how lifestyle and environmental inputs, including diet, stress, toxins, allergens, and microbes, interact with the human body to create imbalances which can commonly lead to chronic health issues.
So, let me take a moment to discuss something. The human body is a system. And this entire system is made up of smaller, �mini-systems� which are all dynamically interacting. But, when one or more of these systems get out of balance, you can become sick. And when these systems get in balance again, you become healthy. And that�s what functional medicine is. Functional medicine is simply understanding what causes imbalances in the human body and treating them to restore balance and provide essential needs to all the systems in the human body. Creating overall health and wellness. It is the science of creating well-being. And doctors achieve this by utilizing a patient�s detailed medical history, combined with targeted lab tests. Doctors who follow a functional medicine approach, like me, by way of instance, generally evaluate your gut and your microbiome, which not many doctors look into. We also want to look at your immune system and whether inflammation is affecting you, something which we refer to as defense and repair. And we want to know how your mitochondria produces energy from food and oxygen. Functional medicine practitioners want to know if you have any dysfunctions in energy production, which is typically the source of numerous diseases, such as Alzheimer�s disease and autism, type 2 diabetes, and fatigue, among many others. We also determine your toxic load and your ability to detoxify, involving the function of other communication systems in your body, such as your hormones. Finally, we evaluate your structural system, from your cells to your biomechanical structures and how these interact with your beliefs, emotions, and more.
I�ve used these procedures, over and over again, to help reverse chronic disease in my patients and educate them on how to achieve long-term health and wellness, all while feeling like the best versions of themselves. We are all given the opportunity to cure or tremendously improve health issues or problems which are often misdiagnosed and mistreated by conventional medicine. Functional medicine provides the opportunity to discover overall well-being at any age.
As a matter of fact, I treated a man named, George, who was 63 years old, weighed 300 pounds, and who had a variety of health problems. He had acid reflux, sinus problems, diabetes, angina heart failure, prostate problems, sexual dysfunction, and edema in his legs. And as a result, he was under a wide array of prescription medicine. He came into my office one day and he said to me, �Can you help me.� I said, �Yes, but you will have to do everything I say.�
So, I recommended him an anti-inflammatory diet, consisting of whole foods and a low intake of sugar, and I advised him to take a variety of supplements in order to optimize his nutrition, all while I guided him into participating on exercises and physical activities. Within a year, it was like a miracle, the patient had lost 155 pounds, he reversed all of his health issues and he stopped using his prescription medicines. Now, he�s planning the rest of his life, when before he came to visit me, he was planning the end of his life. Another patient I treated, named Isabel, struggled with an autoimmune disease. She was only 10 years old and she was already under a pile of drugs, steroids, immunosuppressants, and even chemo drugs. It turns out that her diet and exposure to heavy metals was affecting her gut and causing inflammation. Now, I simply treated the source of her problems and her autoimmune disease went away. She stopped using her prescription medicines and, altogether, her quality of life improved and she thrived.
Functional medicine is an alternative treatment approach which focuses on the interactions between external or environmental factors as well as internal factors associated with the gastrointestinal, endocrine, and immune systems of the human body. Finding the right doctor can make a big difference when it comes to getting health care over sick care. Functional medicine treats the source of the problem rather than treating the symptoms alone. As a chiropractor and functional medicine practitioner, my primary goal is to provide patients with the care they deserve for their health issues as well as to educate them on the fundamentals of functional medicine. The purpose of this article is to help patients find the right doctor and build a healthy doctor-patient relationship. Dr. Alex Jimenez D.C., C.C.S.T. Insight
Sick Care vs Health Care Overview
Over the next few weeks, we�re going to go through some of the fundamental principles and concepts that helped me find the underlying cause of disease through the use of functional medicine. And that will help you achieve overall health and wellness.
In our next article, I�m going to teach you how you can take your own measurements to understand your own well-being and what you can do to improve your vital signs. This procedure is going to help provide you with important baseline data and information to help you better understand your health risks. It�s also going to allow you to evaluate and analyze your own progress as you work towards your ultimate wellness goals.
In the third article, I�m going to try to cover everything about nutrition. I will explain how we can test your nutritional status and how you can utilize food as treatment to start changing your overall health and wellness as soon as possible. Your diet is one of the most effective parts you can control to create a healthier lifestyle. I�ll be sharing tips and tricks for a better nutrition.
Because hormones can impact nearly every aspect of our health, we�re also going to be highlighting them in the fourth article. Unfortunately, a majority of doctors do not understand what optimal hormone levels should look like nor are they aware of the proper methods for testing hormones. I will be preparing you to have an effective conversation with your doctor on which type of hormone test actually matters, what they mean, and what you can do about them.
In article five I�m going to focus on heart health, such as high blood pressure, high cholesterol, and cardiovascular disease. These conditions are extremely common, and unfortunately, conventional medicine frequently tries to treat them by simply controlling the symptoms. Lowering cholesterol, lowering your blood pressure, that does not solve the root cause of the problems. Most of the time, increased levels of blood sugar, obesity and diabetes are the cause of cardiovascular disease. I�m going to demonstrate how we can prevent and reverse these diseases using lifestyle modifications, including nutrition, to improve heart health.
In the sixth article, I�ll be discussing obesity and diabetes, something I�ll be frequently referring to as diabesity. Metabolic disorders ranging from minimal fat and moderate insulin resistance to pre-diabetes to type 2 diabetes. The great thing about these metabolic disorders, however, is that they are completely reversible. But most conventional doctors don�t know it�s reversible. I�m going to help teach you how you can recover a healthy metabolism and how you can reverse the range of problems involved in diabesity, which as you will learn, are a lot of different diseases.
In article seven, I�m going to talk about the immune system, particularly discussing hidden inflammation and disease. As inflammation is described to be the underlying cause of a majority of chronic diseases, we continue to see increasing rates of autoimmune diseases and allergies, which are signs that our immune systems are not functioning properly. However, inflammation is also associated with obesity, type 2 diabetes, heart disease, cancer, dementia, and even depression. I�ll explain how you can talk to your doctor about testing for inflammation, what the markers mean, and how you can care for your immune system on a deeper level.
In article eight, we�re going to move into the topic of the metabolism and mitochondria. Now every one of our cells hold hundreds or thousands of mitochondria, the energy generators of our cells. When these become damaged, we can suffer from a variety of problems, including pain, memory loss, fatigue, and many other symptoms. I�m going to explain how we can care for your mitochondria at the cellular level and why this is a vital piece of optimal health and wellness.
Then, in article nine, we�re going to explore the meaning of detoxification, or the human body�s innate detox system and why this is an essential part of the healing process. From mold to heavy metals, and other toxins, I�m going explain what you need to know to avoid toxins and how you can eliminate those that you are already carrying to optimize your capability to detoxify. With the amount of toxins we�re exposed to on a regular basis, it�s important to identify the causes that may be affecting you and how you can begin eliminating them right away. Fortunately, this is something you can easily do to create a cleaner lifestyle for you and your loved ones.
Finally, in article ten we will be talking about digestion. Digestive health is one of the most commonly discussed topics in functional medicine. Because our digestive system is the center of our health, by diagnosing problems in this system, we can improve everything from mental health, nutrient absorption, and cardiovascular risk to immune system function. In the tenth article, I�m going to guide you through the tests that are available for monitoring your gut health, as well as what you can do to improve your own digestive health, as soon as possible.
I�m so happy you�ll be joining me throughout this series of articles because the future of medicine depends on it. When you learn how you can impact your well-being by being proactive and by creating a partnership with your doctor, everything changes. Through functional medicine, you can also help transform our sick care system into an actual health care system.
So, let�s finish up this article with the questions you want to ask your doctor as you interview them to see if they can be a partner for you and your overall health and wellness. These are just a few high-level questions to start with your discussion. Are you willing to work with me as a partner for my well-being? Also, what�s your point of view on nutrition and health? Do you think food is medicine? Are you willing to give me copies of my test results and explain what they mean? The reason why we�re asking these questions is to make sure that your doctor is willing to work with you and understand the role of functional medicine.
I�m so happy to be a part of your journey to better health and wellness. Thank you very much for joining me today. The scope of our information is limited to chiropractic and spinal health issues as well as functional medicine topics and discussions. To further discuss the subject matter, please feel free to ask Dr. Alex Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topic Discussion:�Acute Back Pain
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief. �
Why is it that the ketogenic diet and intermittent fasting always seem to fall within the same topic of conversation? This is simply because intermittent fasting may be utilized as an instrument to achieve ketosis, the metabolic state associated with the keto diet. During intermittent fasting, the human body is depleted of glycogen stores. Once these glycogen stores are eliminated, fat stores are then released into the bloodstream in order to be converted into energy molecules, known as ketones, from the liver.
What is Ketosis?
Ketosis is a metabolic state which uses ketone bodies, or ketones, as fuel for energy. On a normal carbohydrate-based diet, the human body burns glucose as its main fuel source, where excess glucose is subsequently stored as glycogen. If the human body cannot utilize sugar as fuel for energy, it will utilize glycogen as fuel for energy. Once glycogen is depleted, you begin to burn fat. The ketogenic diet generates a metabolic state which enables you to break down fat into ketones, or ketone bodies, in the liver for energy.
There are 3 major types ketone bodies found in the blood, urine, and breath, including:
Acetoacetate: The type of ketone which is created first. It may be converted to beta-hydroxybutyrate or flipped into acetone.
Acetone: Made spontaneously in the breakdown of acetoacetate. It is a very volatile ketone and it is frequently detectable on the breath once an individual first enters ketosis.
Beta-hydroxybutyrate (BHB): The type of ketone which is utilized for energy and is most abundant on the bloodstream as soon as you’re completely into ketosis. It is the kind that is located in exogenous ketones and what blood tests quantify.
Intermittent Fasting in the Keto Diet
Intermittent fasting is composed of eating within a specific feeding window rather than eating throughout the day. Each individual, whether they are conscious of it or not, fasts intermittently from dinner to breakfast. There are lots of methods to intermittent fasting. A few individuals fast for 16-20 hours intervals on alternate days while others follow a 24-hour day fast. The most common intermittent fasting variety is the 16/8 method, in which you eat in an 8-hour window followed by a 16-hour fasting window.
Other fasting programs incorporate the 20/4 or even 14/10 methods. Other people follow 24-hour fasts one or two times each week. Intermittent fasting can get you in ketosis quicker because your cells will immediately absorb your glycogen stores and begin burning fat. However, what about once you get into ketosis? Is intermittent fasting worth following consistently? Following the ketogenic diet and intermittent fasting can be a great addition towards an individual’s overall health and wellness, providing various health benefits.
The keto diet and intermittent fasting can provide the following health benefits, including:
Healthy weight-loss
Fat reduction, not muscle reduction
Balancing cholesterol levels
Enhancing insulin sensitivity
Maintaining blood glucose levels steady
Health Benefits of the Ketogenic Diet
The ketogenic diet dramatically reduces your caloric intake, forcing your body to burn fat instead of sugar, which makes it a powerful tool for weight reduction. While individual results vary, the keto diet has always resulted in a decrease in body fat in a selection of situations. Within a 2017 study, subjects who followed a very low carbohydrate keto meal program significantly decreased body fat percentage and body fat mass, losing an average of 7.6 lbs and 2.6 percent body fat while preserving lean muscle mass.
Likewise, a 2004 research detecting the long-term consequences of a ketogenic diet in overweight patients discovered that the weight and body mass of those patients diminished dramatically over the span of two decades. Individuals who radically reduced their carb intake saw a substantial decline in LDL (bad) cholesterol, triglycerides, and enhanced insulin sensitivity. In 2012, researchers compared a ketogenic diet to eating fewer calories for overweight kids and adults. The results showed kids after the keto diet lost significantly more body fat. They also revealed a dramatic decline in insulin levels, a biomarker of Type 2 diabetes.
Health Benefits of Intermittent Fasting
Studies have shown that intermittent fasting may be an effective weight loss tool, more powerful than just cutting calories. In one analysis, intermittent fasting has been proven to be as successful as constant calorie restriction in combating obesity. In studies done by the NIH, there was reported weight reduction with over 84 percent of participants, regardless of which fasting program they picked.
Much like ketosis, intermittent fasting increases fat loss while preserving lean muscle mass. In one study, researchers reasoned that fasting led to greater weight loss compared to a low-carb diet, though the overall caloric consumption was exactly the same. If you are attempting to lose weight, then a keto diet or intermittent fasting can be a massive help. But that is not where the rewards stop.
Intermittent Fasting and the Keto Diet for Mental Health
Both intermittent fasting and the ketogenic diet can provide various mental health advantages. Both have been clinically shown to boost memory, improve mental clarity and focus, as well as prevent the development of neurological disorders like Alzheimer’s and epilepsy. On a carb-based diet, changes in glucose can cause changes in energy levels. During ketosis, your brain employs a more consistent supply of fuel: ketones from the fat stores, leading to better productivity and psychological performance.
Whenever you’ve got a consistent and clean energy source from ketones, the brain works better. In addition to this, ketones are better at protecting your brain. Studies reveal that ketone bodies might have antioxidant properties which protect your brain cells from free radicals and oxidative stress. In one study conducted on adults with diminished memory, the growth of BHB ketones in their own blood helped enhance cognition. Also, when you’ve got difficulty staying focused, your hormones can be to blame.
Your brain has two chief neurotransmitters: glutamate and GABA. Glutamate will help you form new memories, and get your brain cells to communicate with one another. GABA is what helps restrain glutamate. If there is too much glutamate, it can cause brain cells to quit working and finally perish. GABA is there to control and slow down glutamate. If GABA levels are reduced, glutamate reigns free and you experience mental fog. Ketones stop damage to cells by processing surplus glutamate into GABA. Considering that ketones raise GABA and lessen glutamate, they assist in preventing cell damage, preventing cell death and enhancing mental focus.
Researchers believe that intermittent fasting enhances memory, decreases oxidative stress, and conserves learning abilities. Since your cells are under moderate strain whilst fasting, the top cells adapt to the stress by improving their particular ability to deal with these circumstances while the weakest tissues die. This is much like the strain that your body gets when you reach the gym.
Exercise is a kind of stress that your body adjusts to improve and get more powerful. This also applies for intermittent fasting: so long as you are still alternate between routine eating habits and fasting, it is going to continue to benefit you. Implying equally that ketosis and intermittent fasting will help improve your cognitive functioning because of the synergistic and protective effects of ketones.
The ketogenic diet and intermittent fasting are two different nutritional strategies which provide many common health benefits. According to various research studies, both the keto diet and intermittent fasting can help boost ketones, helping the body burn fat more efficiently than any other nutritional strategy. And when these are utilized together, they definitely form a powerful dietary program. The article above discusses the differences between the ketogenic diet and intermittent fasting as well as demonstrates the health benefits of both of these dietary programs and how they can help improve overall health and wellness. Dr. Alex Jimenez D.C., C.C.S.T. Insight
The Perks of Intermittent Fasting and the Keto Diet
The ketogenic diet and intermittent fasting possess similar health benefits because both approaches involve ketosis. Ketosis has lots of physical and mental advantages, from weight loss to enhanced brain function. People following a ketogenic diet may use intermittent fasting as a tool to achieve ketosis and enhance their general well-being. The scope of our information is limited to chiropractic and spinal health issues. To discuss the subject matter, please feel free to ask Dr. Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topic Discussion:�Acute Back Pain
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief. �
If you are currently thinking about the ketogenic diet, then you might be asking yourself, is the keto diet right for you? While you may have already heard about the benefits of the ketogenic diet, you might still be wondering about whether if it is worth it to completely change your diet to take advantage of these benefits.
The keto diet has many benefits, from weight loss and improved physical health to mental clarity and enhanced physical performance. In the following article, we will dive into the details of some of the ketogenic diet health benefits. These benefits can help with the particular health goal you may be attempting to attain.
Ketogenic Diet and Weight Loss
In comparison to low-fat dieting, a low-carb diet can deliver superior results within a shorter time period in terms of weight loss, and the management of cholesterol, and blood pressure. If you want to shed weight, the ketogenic diet plan provides the following benefits and will get you closer to attaining your objective. There can be many reasons for this, including:
Low-carb and ketogenic diets are more satisfying with their low carb content and higher quantities of fats and protein.
Going onto a low-carb diet usually makes you lose extra water weight.
Most individuals can undergo weight loss fairly quickly, especially within the first week�of beginning a ketogenic diet.
Increased HDL Cholesterol
Together with the high consumption of saturated fats and other healthy fats, the ketogenic diet may help raise HDL cholesterol and enhance triglycerides levels. Both of these are�considerably significant towards promoting heart health.
Ketogenic Diet and Physical Health
Acne
Following the ketogenic diet has been demonstrated to also be able to help reduce inflammation and lesions of the skin like those found in acne. This is believed to occur due to the effects of ketosis, or the state in which the cells use ketones instead of glucose for energy.
IBS Support
Moreover, several research studies have also associated a link between the reduced consumption of glucose, or sugar, and an improvement in symptoms of irritable bowel syndrome, or IBS. As a matter of fact, one research study demonstrated that following a ketogenic diet may improve bowel movement habits and help reduce abdominal pain, improving quality of life in people with IBS.
Ketogenic Diet and Physical Performance
Balanced Energy Levels
Do not be surprised if you’re ready to stop drinking coffee every day after adapting to the keto diet. Achieving and maintaining ketosis involves benefits like no day slumps, no mood swings, and reducing changes in energy levels that you might experience otherwise.
In addition, you’ll likely find it much easier to remain longer periods of time without feeling hungry. This is what ultimately helps with weight loss, steady blood sugar levels, and extended periods of fasting, which is one of the best ways to get into ketosis.
Enhanced Workouts
Adjusting to the ketogenic diet may take time, however, once your body gets used to burning fat for fuel rather than sugar, or glucose, from carbohydrates, you will likely notice a difference in your physical performance and endurance, such as more energy and focus for workouts. This makes sense because being in ketosis “instructs” the entire human body to burn fat for fuel more efficiently.
The most important first step in case you start the ketogenic diet and notice limitations in your physical performance is to give your body some time to adapt from utilizing carbohydrates as its primary fuel to utilizing ketones as a source of energy. For individuals who participate in a lot of physical activities and exercise as well as athletes may benefit from a cyclical or targeted ketogenic diet.
Fat Loss / Muscle Gain
The amount of protein intake on a ketogenic diet makes it excellent for building muscle mass. Results might seem to come more gradually than for someone fueling their workouts but that is usually because you’re building lean mass together with fat reduction. By way of instance, when documenting a keto fast for four days, the individual gained 2.4 lbs of muscle with 1.1 lbs of fat reduction.
Ketogenic Diet and Mental Clarity
Several research�studies have demonstrated that a ketogenic diet may have the ability to support mental clariy as well as help boost productivity, support better memory, and also, have positive effects in regard to moderate cognitive impairment.
Neurological Support
Early usage of the ketogenic diet has been used as a treatment for reducing seizures in people with epilepsy, especially children. Additionally, it has been shown to benefit people with Parkinson’s disease, Alzheimer’s disease, and other neurodegenerative disorders. This is likely because ketone bodies created through the keto diet can have neuroprotective effects.
Weight loss is one of the most well-known advantages of the ketogenic diet, however, this nutritional plan can have many other health benefits. By reducing the consumption of carbohydrates, the cells will go into a state of ketosis and instead utilize ketones created from fats, providing a steadier supply of energy than that of glucose, or sugar. Furthermore, research studies have also demonstrated the ketogenic diet’s possible role in disease prevention, such as for people with epilepsy. Dr. Alex Jimenez D.C., C.C.S.T. Insight
The benefits of the ketogenic diet are essential, not just for weight loss, but for overall health and wellness. When you are eating more fats and proteins with fewer carbohydrates, you are more likely to end up eating fewer calories. With this, you also don’t experience a change of energy levels but instead maintain a level of energy that lets you remain focused on your everyday tasks.
Regardless of the health goal you have in mind, the ketogenic, or keto, offers many benefits to improve your quality of life. Being aware of the proper foods you should eat on the keto diet is also important. The scope of our information is limited to chiropractic and spinal health issues. To discuss the subject matter, please feel free to ask Dr. Jimenez or contact us at�915-850-0900�.
Curated by Dr. Alex Jimenez
Additional Topic Discussion:�Acute Back Pain
Back pain�is one of the most prevalent causes of disability and missed days at work worldwide. Back pain attributes to the second most common reason for doctor office visits, outnumbered only by upper-respiratory infections. Approximately 80 percent of the population will experience back pain at least once throughout their life. The spine is a complex structure made up of bones, joints, ligaments, and muscles, among other soft tissues. Injuries and/or aggravated conditions, such as�herniated discs, can eventually lead to symptoms of back pain. Sports injuries or automobile accident injuries are often the most frequent cause of back pain, however, sometimes the simplest of movements can have painful results. Fortunately, alternative treatment options, such as chiropractic care, can help ease back pain through the use of spinal adjustments and manual manipulations, ultimately improving pain relief. �
IFM's Find A Practitioner tool is the largest referral network in Functional Medicine, created to help patients locate Functional Medicine practitioners anywhere in the world. IFM Certified Practitioners are listed first in the search results, given their extensive education in Functional Medicine